The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries.
David Lubin Memorial Library Cataloguing in Publication Data
FAO, Rome (Italy)
Maize in human nutrition.
(FAO Food and Nutrition Series, No. 25)
ISBN 92-5-103013-8
1. Maize
2. Human Nutrition
I. Title
II. SeriesFAO code: 80
AGRIS: SO1
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying or otherwise, without the prior permission of the copyright owner. Applications for such permission, with a statement of the purpose and extent of the reproduction, should be addressed to the Director, Publications Division, Food and Agriculture Organization of the United Nations, Viale delle Terme di Caracalla, 00100 Rome, Italy.
(c) FAO 1992
Over the years FAO has published a series of nutrition studies. Maize and maize diets, a nutritional survey published in 1953, covered available information and knowledge on maize up to that date. Since then a vast amount of research information on breeding, varietal improvement' storage, processing, consumption and improvement of the nutritional quality of maize has become available.
The need to update and revise the old edition to include this information was keenly felt by FAO, which has decided to publish it under a new title, Maize in human nutrition, and to aim it at a more technical level of readership.
The current edition provides expanded information on the chemical composition of maize, including the makeup of maize protein and dietary fibre, on grain quality and storage and on the effects of lime-cooking of maize and the manufacture of foods such as tortillas, arepas and ogi. It reviews evidence of the association between maize consumption, bound niacin and pellagra and presents the evidence of amino acid deficiencies in maize and results obtained from experiments with both humans and animals. It discusses the importance of raising the protein quality of maize through incorporation of the opaque-2 gene and its probable contribution to improving the diet of maize-eating populations, and it makes a strong case for commercial production of quality protein maize (QPM). Finally, it provides a more up-to-date account of how maize diets can be improved following the old principle of nutrition: consume a balanced diet containing food legumes, animal protein, fruits and vegetables.
FAO acknowledges the contribution of Prof. R. Bressani of the Institute of Nutrition of Central America and Panama for the extensive revision and rewriting of the book with the technical assistance of Ms Maria Antonietta Rottman. Dr M.A. Hussain, officer-in-charge of Nutrition Programmes Service, Food Policy and Nutrition Division, was responsible for technical editing and preparation of the final manuscript. Valuable suggestions were made by other staff members of the division and of the Plant Production and Protection Division and Agricultural Services Division.
This book is intended for nutritionists, agronomists, food scientists, dieticians and others concerned with maize. It is hoped that they will find it useful and worthwhile.
Paul Lunven
Director
Food Policy and Nutrition Division
Maize, the American Indian word for corn, means literally "that which sustains life". It is, after wheat and rice, the most important cereal grain in the world, providing nutrients for humans and animals and serving as a basic raw material for the production of starch, oil and protein, alcoholic beverages, food sweeteners and, more recently, fuel. The green plant, made into silage, has been used with much success in the dairy and beef industries. After harvest of the grain, the dried leaves and upper part, including the flowers, are still used today to provide relatively good forage for ruminant animals owned by many small farmers in developing countries. The erect stalks, which in some varieties are strong, have been used as long-lasting fences and walls.
Botanically, maize (Zea mays) belongs to the grass family (Gramineae) and is a tall annual plant with an extensive fibrous root system. It is a cross pollinating species, with the female (ear) and male (tassel) flowers in separate places on the plant. The grain develops in the ears, or cobs, often one on each stalk; each ear has about 300 to 1 000 kernels, weighing between 190 and 300 g per I 000 kernels, in a variable number of rows (12 to 16). Weight depends on genetic, environmental and cultural practices. Grain makes up about 42 percent of the dry weight of the plant. The kernels are often white or yellow in colour, although black, red and a mixture of colours are also found. There are a number of grain types, distinguished by differences in the chemical compounds deposited or stored in the kernel.
Special crops grown primarily for food include sweet corn and popcorn, although dent, starchy or floury and flint maize are also widely used as food. Flint maize is also used as feed. Immature ordinary corn on the cob either boiled or roasted is widely consumed. Floury maize is a grain with a soft endosperm much used as food in Mexico, Guatemala and the Andean countries. The dent type of maize has a vitreous horny endosperm at the sides and back of the kernel, while the central core is soft. Flint kernels have a thick, hard and vitreous endosperm surrounding a small, granular, starchy centre.
The cultivation of maize or Indian corn most probably originated in Central America, particularly in Mexico, from whence it spread northward to Canada and southward to Argentina. The oldest maize, about 7 000 years old, was found by archaeologists in Teotihuacan, a valley near Puebla in Mexico, but it is possible that there were other secondary centres of origin in the Americas. Maize was an essential item in Mayan and Aztec civilizations and had an important role in their religious beliefs, festivities and nutrition. They claimed that flesh and blood were made from maize. The survival of the oldest maize and its distribution depended on humans who harvested the seed for the following planting. At the end of the fifteenth century, after the discovery of the American continent by Christopher Columbus, maize was introduced into Europe through Spain. It then spread through the warmer climates of the Mediterranean and later to northern Europe. Mangelsdorf and Reeves (1939) have shown that maize is grown in every suitable agricultural region of the world and that a crop of maize is being harvested somewhere around the globe every month of the year. Maize grows from latitude 58° in Canada and the former Union of Soviet Socialist Republics to latitude 40° in the Southern Hemisphere. Maize crops are harvested in regions below sea-level in the Caspian Plain and at altitudes of more than 4 000 m in the Peruvian Andes.
In spite of its great diversity of form, all main types of maize known today were apparently already being produced by the native populations when the American continent was discovered. All maize is classified as Zea mays. Furthermore, evidence from botany, genetics and cytology has pointed to a common origin for every existing type of maize. Most researchers believe that maize developed from teosinte, Euchlaena mexicana Schrod, an annual crop that is possibly its closest relative. Others, however, believe that maize originated in a wild maize that is now extinct. The closeness of teosinte to maize is suggested by the fact that both have ten chromosomes and are homologous or partially homologous.
Introgression between teosinte and maize has taken place in the past and still does today in areas of Mexico and Guatemala where teosinte grows among the maize crop. Galinat (1977) indicated that of the various hypotheses on the origin of maize, essentially two alternatives remain viable: first, that present-day teosinte is the wild ancestor of maize and/or that a primitive teosinte is the common wild ancestor of both maize and teosinte or, second, that an extinct form of pod maize was the ancestor of maize, with teosinte being a mutant form of this pod maize.
In any case, most of the modern varieties of maize have been derived from materials developed in the southern United States of America, Mexico and Central and South America.
The maize plant may be defined as a metabolic system whose end product is mainly starch deposited in specialized organs, the maize kernels.
The development of the plant may be divided into two physiological stages. In the first or the vegetative stage, different tissues develop and differentiate until the flower structures appear. The vegetative stage is made up of two cycles. In the first cycle the first leaves are formed and development is upward. Dry matter production in this cycle is slow. It ends with the tissue differentiation of the reproductive organs. In the second cycle the leaves and reproductive organs develop. This cycle ends with the emission of the stigmas.
The second stage, also known as the reproductive stage, begins with the fertilization of the female structures, which will develop into ears and grains. The initial phase of this stage is characterized by an increase in the weight of leaves and other flower parts. During the second phase, the weight of the kernels rapidly increases (Tanaka and Yamaguchi, 1972).
The plant develops morphological characteristics and differences in the vegetative and reproductive stages as evolutionary consequences of natural selection and domestication. Some genotypes have adapted to specific ecological zones and so have developed such barriers as day-length sensitivity and temperature sensitivity, which limit their adaptability to specific areas of latitude and altitude. Thus improvement programmes must be conducted within the areas where the improved varieties are to be grown. This does not mean, however, that specific genetic characteristics can be attained by backcrossing.
The morphology or architecture of the plant has also suffered evolutionary pressures that have resulted in great variability in the number, length and width of leaves, plant height, positions of ears, number of ears per plant, maturation cycles, grain types and number of rows of grain, among many other characteristics.
This variability is of great value in improving the productivity of the plant and specific organic components of the grain. The main yield components include the number and weight of grains. These yield components are determined by quantitative genetic effects that can be selected relatively easily. The number of grains depends on the ear and is determined by the number of rows and the number of kernels per row. The size and shape of the kernel determine its weight in the presence of other constant factors such as grain texture and grain density. The ratio of grain weight to total plant weight for most maize lines is about 0.52. From 100 kg of cobs, about 18 kg of grain is obtained. One ha of maize yields about 1.55 tonnes of stalk residue. In field-dried maize plants from three locations in Guatemala, plant dry weight varied from 220 to 314 g. This weight comprised 1.8 percent dried flowers, 14.7 to 27.8 percent stalks, 7.4 to 15.9 percent leaves, 11.7 to 13 percent husks, and 9.7 to 11.5 percent cobs. The field-dried grain represented 30 to 55.9 percent of the whole plant dry weight. These data show the significant yield of plant residues that are often left in the field. The distribution may change, however, since it is accepted that about half of the dry matter is grain and the other half is made up of plant residues excluding roots (Barber, 1979).
Maize kernels develop through accumulation of the products of photosynthesis, root absorption and metabolism of the maize plant on the female inflorescence called the ear. This structure may hold from 300 to 1 000 single kernels depending on the number of rows, diameter and length of the cob. Kernel weight may be quite variable, ranging from about 19 to 40 g per 100 kernels. During harvest the ears of maize are removed from the maize plant either by hand or mechanically. The husks covering the ear are first stripped off, then the kernels are separated by hand or, more often, by machine.
TABLE 1 - Weight distribution of main parts of the kernel
| Structure | Percent weight distribution |
| Pericarp | 5-6 |
| Aleurone | 2-3 |
| Endosperm | 80-85 |
| Germ | 10-12 |
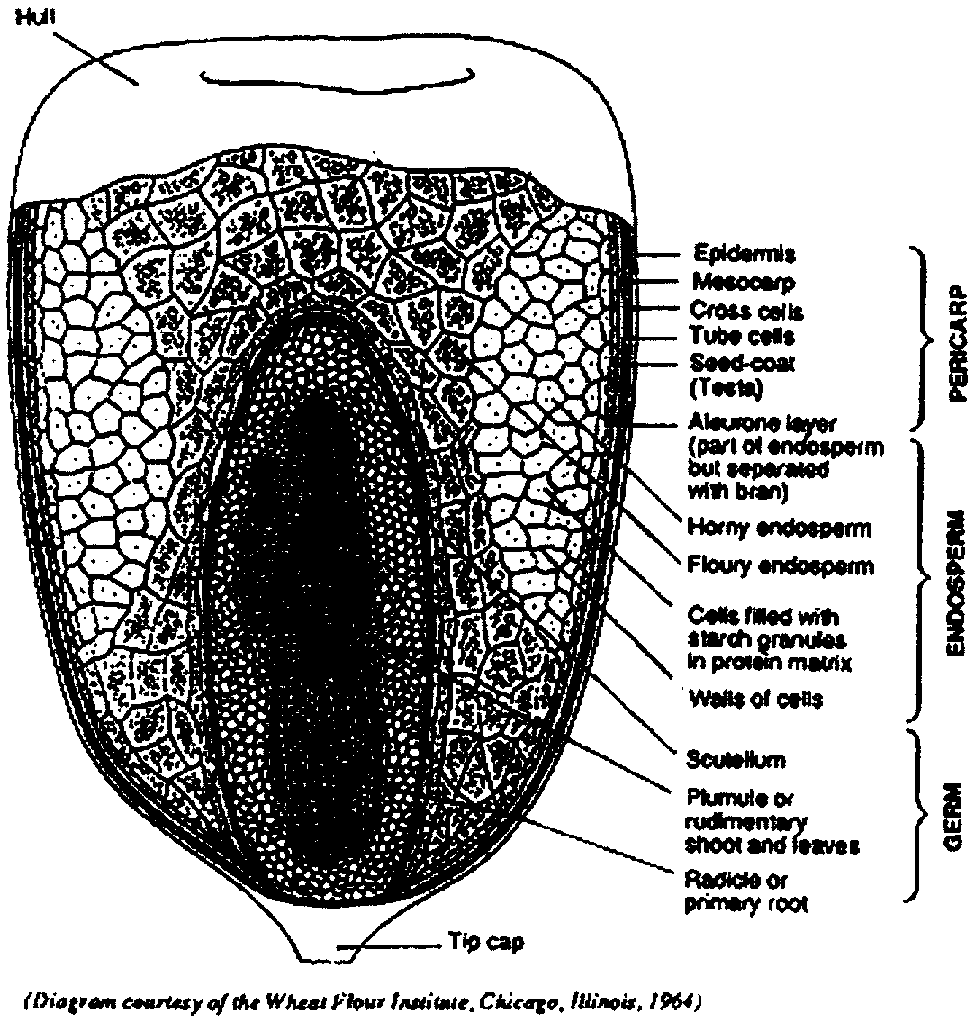
The maize kernel is known botanically as a caryopsis; a single grain contains the seed coat and the seed, as shown in Figure 1. The figure also shows the four major physical structures of the kernel: the pericarp, hull or bran; the germ or embryo; the endosperm; and the tip cap (dead tissue found where the kernel joins the cob). The gross anatomy and the microscopic structure of these anatomical components were well described by Wolf et al. (1952) and by Wolf, Khoo and Seckinger (1969). They also studied the structure of the improved opaque-2 maize and found differences between its endosperm and that of common maize. The protein matrix was thinner and there were fewer and smaller protein bodies, since there is a restriction in zein synthesis in opaque-2 maize. Robutti, Hoseny and Deyoe (1974) and Robutti, Hoseny and Wasson (1974) reported on the protein distribution, amino acid content and endosperm structure of opaque-2 maize.
The weight distribution of the different parts of the maize kernel is shown in Table 1. The endosperm, the lqrgest structure, provides about 83 percent of the kernel weight, while the germ averages 11 percent and the pericarp 5 percent. The remainder is the tip cap, a conical structure that together with the pedicel attaches the kernel to the ear of maize. Table 2 shows the distribution of weight and nitrogen among the anatomical parts of common and selected kernel varieties, such as high-oil and high-protein maize and three quality protein maize (QPM) selections (Bressani and Mertz, 1958). The main difference in the high-oil variety is the size of the germ, which is about three times as large as the germ from common maize with a reduction in endosperm weight. Germ of the high-protein varieties is larger than that of common maize but about half the size of high-oil varieties. There are also differences in the weight of the seed-coats. Table 2 also shows some data for teosinte, the closest relative to maize. The seed weight is much lower than that of maize seed, and the endosperm weighs about half that of maize. The three QPM selections are similar to maize in weight per seed and in weight of the seed-coat, the endosperm and the germ. Similar data have been reported by other authors. Table 3 summarizes data for two common varieties and one opaque-2 maize (Landry and Moureaux, 1980). The two common samples have the same general characteristics as those reported above; the opaque-2 sample, however, has a larger germ providing more nitrogen than the QPM selections in Table 2. With respect to the germ, the increase of weight and of nitrogen amounts in absolute as well as relative terms is consistent with other results (Watson, 1987).
FIGURE 1 - Maize kernel structure: longitudinal section enlarged approximately 30 times
TABLE 2 - Distribution of weight and nitrogen among parts of the kernel
| Maize sample | Weight of 20 seeds (g) | Weight distribution (%) |
Total N (%) | Nitrogen distribution (%) |
||||
| Seedcoata | Endosperm | Germ | Seedcoat | Endosperm | Germ | |||
| US 4251 | 5.62 | 6.3 | 86.3 | 7.4 | 1.31 | 3.3 | 81.2 | 15.5 |
| US high oil (HO) | 5.72 | 6.4 | 71.2 | 22.4 | 1.99 | 2.4 | 68.4 | 29.2 |
| US high protein (H5) | 4.32 | 6.9 | 82.7 | 10.4 | 2.24 | 2.2 | 83.2 | 14.6 |
| US high protein (HP) | 4.97 | 7.4 | 78.9 | 13.7 | 2.14 | 2.7 | 78.2 | 19.1 |
| US normal-Sh1 PT | 4.38 | 6.7 | 79.6 | 13.7 | 2.14 | 2.7 | 78.2 | 19.1 |
| US normal mutant-Sh1 PT | 2.50 | 10.7 | 70.6 | 18.7 | 2.21 | 6.1 | 64.6 | 29.3 |
| Tiquisate (TGY)(Guat.) | 8.24 | 4.9 | 83.9 | 11.2 | 1.37 | 2.8 | 75.2 | 22.0 |
| San Sebastian (SSD)(Guat.) | 8.24 | 4.9 | 83.9 | 11.2 | 1.37 | 2.8 | 75.2 | 22.0 |
| Guatemalan 142-48 | 6.91 | 6.9 | 82.1 | 11.0 | 1.83 | 2.6 | 81.0 | 16.4 |
| Guatemalan Cuyuta | 5.95 | 5.7 | 82.5 | 11.8 | 1.28 | 2.9 | 72.4 | 24.7 |
| Guatemalan teosinte | 1.56 | 55.6b | 44.4 | - | 1.81c | 8.2 | 91.8d |
|
| Nutricta QPM | 5.91 | 5.7 | 82.7 | 11.6 | 1.42 | 1.7 | 72.8 | 25.5 |
| QPM yellow | 6.49 | 5.9 | 81.6 | 12.5 | 1.48 | 2.4 | 73.4 | 24.2 |
| QPM white | 5.31 | 5.9 | 82.4 | 1.6 | 1.36 | 1.4 | 72.8 | 25.7 |
aPericarp plus tip cap
bIncludes the seed-coat (1.3%) and the hull (54.3%)
cThe hull contained 0.26% nitrogen; dehulled teosinte
contained 3.81 % nitrogen
dIncludes the germ
Source: Bressani and Mertz, 1958
TABLE 3 - Weight and nitrogen distribution of parts of common and opaque-2 maize kernels
| Part of kernel | Dry matter (%) |
Nitrogen (%) |
||||
| Common | Common | Opaque-2 | Common | Common | Opaque-2 | |
| Germ | 13.5 | 8.1 | 35 | 20.1 | 14.9 | 35.1 |
| Endosperm | 80.0 | 84.0 | 61 | 76.5 | 80.5 | 60.7 |
| Seed coat | 6.5 | 7.9 | 4 | 3.4 | 4.6 | 4.2 |
Source: Landry and Moureaux, 1980
{kind=link}