Chapter 2 Physiological deterioration in cassava: biochemistry of the processes involved
The rapid development of primary or physiological deterioration in cassava has been strongly associated with mechanical damage which occurs during harvesting and handling operations (Booth 1976). Frequently the tips are broken off as the roots are pulled from the ground (Figure 5) and severance from the plant necessarily creates a further wound. In addition, transport from the field to the markets can result in further abrasion. In most cases physiological deterioration develops from sites of tissue damage and is initially observed as blue-black discoloration of the vascular tissue which is often referred to as vascular streaking (Figure 4). Initial symptoms are rapidly followed by a more general discoloration of the storage parenchyma (Figure 6).
In most plants, tissue damage results in a cascade of wound responses (Bowles, 1990) that quickly result in the defence of the wounded tissue and the subsequent sealing of exposed tissue by regeneration of a protective barrier (periderm formation). Common wound responses directly involved in defence include Iytic enzymes (glucanase and chitinase), protease inhibitor proteins and hydroxyproline-rich glycoproteins production. Enzymes associated with the phenylpropanoid pathway, such as phenylalanine ammonia-lyase and chalcone synthase, lead to the biosynthesis of phenolics which may act directly as defence compounds (quinones, phytoalexins) or can form polymers, such as lignin, that render cell walls more resistant to water loss and attack from microbial enzymes.
Cassava roots when stored at high relative humidities (RH) of around 80 to 90 percent show a typical wound-healing response (Figure 7) with periderm formation occurring in seven to nine days at 35 �C and 10 to 14 days at 25�C (Rickard, 1985). This response is notably slower than in the other tropical root crops, such as yam, which form a periderm in four to five days at 35�C (Passam, Read and Rickard, 1976). Booth (1976), however, demonstrated that periderm formation in cassava roots occurred around small v-shaped cuts within four to seven days at 35�C, indicating that the magnitude of the wound sustained can affect the time required for periderm formation. In cassava the formation of a wound periderm (curing) has been found to suppress the development of physiological deterioration (Booth, 1976).
Cytochemical investigations of changes occurring at the wound surface of cut cassava held at high storage RH have shown the development of a number of common plant wound responses (Rickard, 1982; 1985). Along with the development of colourless and coloured deposits at the wound surface and in the underlying cell layers (Figure 7) associated increases were observed in responses to cytochemical tests for phenols, lipids. carbohydrates and lignin as well as in the activity of polyphenol oxidase and peroxidase. Additional changes were also observed using ultraviolet light with the development of whitish-blue fluorescence in the storage parenchyma. The cytochemical and general stains used were not, however, sufficiently specific to determine the exact identity of the material formed at the wound surfaces. However, the phenol test responses indicated the presence of flavanols (catechins and proanthocyanidins). The existence of lignin in the deposits which formed a barrier across the wound surface was not substantiated by either fluorescence or polarized light microscopy, indicating a polyphenolic rather than a lignified wound barrier. However, lignification of the cell walls in this area was substantiated by use of these microscopic techniques.
In cassava roots held at low storage humidity (less than 80 percent RH) the responses to injury do not remain localized at wound surfaces and physiological deterioration generally develops throughout the storage tissue within three to four days after harvest. Respiration experiments by Marriott, Been and Perkins (1979) have indicated that the initial development of physiological deterioration is associated with stress induced by water loss from wounds. Injured cassava roots were found to have a higher respiration rate when held under low humidity storage conditions.
Microscopic observations have shown that the initial response to injury at low storage humidity involves the development of colourless deposits in the xylem parenchyma and an increase in storage parenchyma fluorescence. Material formed in the xylem parenchyma was observed to enter subsequently and occlude the xylem vessels (Figure 8) along with the production of tyloses (Figures 9 and 10). The visual symptoms of vascular streaking were found to develop from discoloration of xylem parenchyma and vessel occlusions.
The initial symptoms of vascular streaking are rapidly followed by a more general discoloration of the storage parenchyma. Prior to the appearance of general tissue discoloration, colourless deposits and intense fluorescence were observed to develop in the storage tissue. Increases in the activities of polyphenol oxidase and peroxidase and a decrease in response to free phenols were noted to accompany the appearance of coloured deposits. The material formed in the xylem system and storage parenchyma gave similar cytochemical test responses to those obtained at the wound surface of cured roots. The presence of phenolic compounds during the development of physiological deterioration was also visually followed by the addition of cytochemical reagents to cut root pieces. Surface test responses for flavanols were strongly associated with areas of storage parenchyma discoloration (Figure 11).
The principal phenolic compounds associated with the development of physiological deterioration have been characterized using a variety of biochemical techniques. Phenolic compounds identified in cassava include scopoletin, scopolin, esculin, proanthocyanidins, (+)-catechin and (+) gallocatechin (Rickard, 1981; 1985;Tanakaetal., 1983;Wheatley, 1982; Wheatley and Schwabe, 1985). Maximum scopoletin content, which is mainly responsible for the strong storage parenchyma fluorescence, was found to peak within 24 hours of injury (Figure 12) and prior to the development of the visual symptoms of physiological deterioration (Wheatley and Schwabe, 1985). Increases in flavanol and proanthocyanidin content continued to increase with the development of physiological deterioration (Rickard, 1981; 1985). Fluorescence microscopy and light microscope cytochemical techniques have demonstrated that, during the development of physiological deterioration, scopoletin and flavanols can be formed throughout the storage parenchyma of cassava and do not involve the activity of specialized cells.
The phenylpropanoid pathway is responsible for the biosynthesis of phenolic compounds (Figure 13) and it has been the subject of much research in different plant systems. The phenylpropanoid pathway is involved in many aspects of wound response including the production of isoflavonoid phytoalexins, flavonoid pigments, ultraviolet protectants and the generation of lignin and suberin (Hahlbrock and Scheel, 1989). Phenylalanine ammonialyase controls the key initial entry into the phenylpropanoid pathway and genes for this and other associated enzymes can begin transcription within two to three minutes of wounding or elicitor treatment (Lorenzo et al., 1987; Fritzemeier et al., 1987). Two enzymes from this pathway, phenylalanine ammonia-lyase and 4coumarate: CoA ligase, have been shown to be differentially induced by wounding (Lois and Hahlbrock, 1992).
The activity of phenylalanine ammonia-lyase was found to increase rapidly in cassava roots along with the development of physiological deterioration (Rickard, 1982; 1985; Tanaka et al., 1983). Peak activity of phenylalanine ammonia-lyase generally occurred some two to three days after injury (Figure 14). The changes in phenolic content together with the enhanced activity of phenylalanine ammonia-lyase observed in cassava indicate that increases in phenolic compounds are at least partly due to de novo synthesis.
In common plant wound responses, phenolic compounds from the phenylpropanoid pathway are acted upon by a range of peroxidases to produce wound-repair substances in the immediate vicinity of the site of tissue damage. The enzymes involved in these processes are synthesized in response to wounding after a time lag which is dependent on the nature of the stimulus. The walls of cells immediately beneath exposed wounds typically become impregnated with phloroglucin-positive material such as lignin and/or suberin and periderm formation occurs beneath this layer (Beeching et al., in press).
In cassava, cytochemical tests for peroxidase gave increased staining responses during the development of a localized wound response (curing) and during the later stages of physiological deterioration (Rickard, 1982; 1985). Biochemical analysis of peroxidase in wounded cassava roots showed that increased activity occurred after a lag period of about one day and continued to increase with the development of physiological deterioration (Tanaka et al., 1983). The electrophoretic patterns of extracts from discolored cassava showed increases in the number and the staining intensities of the soluble, tonically-bound and covalently-bound bands of peroxidases activity. Polyphenol oxidase activity was also found to increase in discolored cassava tissue but, only in the covalently-bound fraction (Plumbley, Hughes and Marriott, 1981).
FIGURE 12 Time course of changes in the amounts of coumarin components in cassava (cv. Golden Yellow) in response to cut injury. Tissues disks were 2mm thick.
{kind=link}
FIGURE 13 Outline of the pathway of phenylpropanoid biosynthesis
{kind=link}
Source: Dr M. Rhodes, AFRC Institute of Food Research, private communication.
FIGURE 14 Timecourse of changes in activities of acid invertase, phenylalanine ammonia-lyase and peroxidase in cassava (cv. Golden Yellow) in response to cut injury. Tissue disks were 2mm thick.
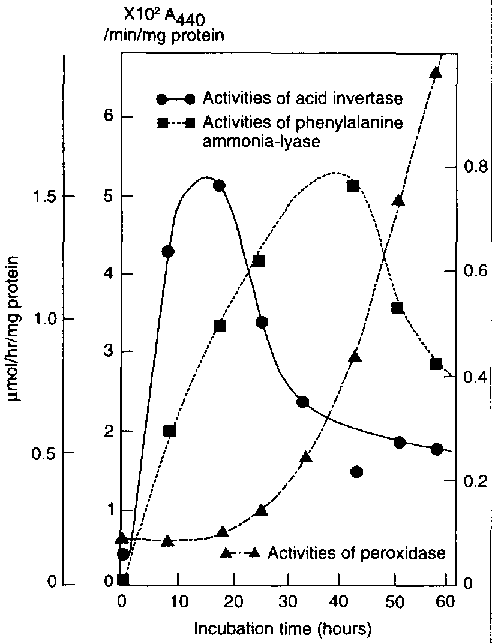{kind=link}
Source: Tanaka et al.,1983.
Activation of genes for enzymes of the phenylpropanoid pathway tends to be localized and occurs only weakly at a distance from the site of excitation (Bowles, 1990). An increasing number of substances are being reported to be capable of acting as wound signals. Volatile substances, such as ethylene or the recently discovered jasmonic acid, have been shown to be important elicitors of wound and defence responses (Ecker and Davis, 1987; Farmer, Johnson and Ryan, 1992).
The development of physiological deterioration throughout the storage tissues of cassava root after harvesting suggests the transmission of intercellular signals from the sites of damage. However, this topic has not been studied in cassava except for the production of ethylene. Like most other plant tissues (Hyodo, 1991), cassava has been found to produce ethylene in response to wounding. Ethylene production from damaged cassava roots was reported to occur after a lag period of about six hours and continued to increase over a 22hour period (Plumbley, Hughes and Marriott, 1981). Similar results were obtained by Hirose, Data and Quevedo ( 1984) after a lag phase of 16 hours with varietal differences affecting the rate of ethylene production (Figure 15). Experimental evidence to date suggests that ethylene is not directly involved in the development of physiological deterioration. Preharvest pruning, which is effective in suppressing physiological deterioration, had no significant influence on ethylene production following injury and the application of endogenous ethylene was not found to affect wound responses (Hirose, Data and Quevedo, 1984).
FIGURE 15 Varietal difference in ethylene production
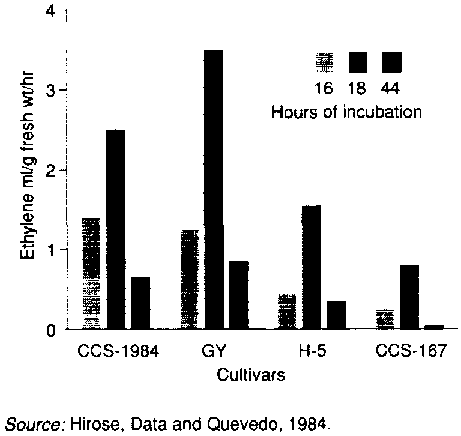{kind=link}
Source: Hirose. Data and Quevedo, 1984.
The maintenance of compartmentation and selective permeability are essential features of membrane function. A number of signal compounds, such as traumatic and jasmonic acid, can be formed from free fatty acids released from the enzymic breakdown of membrane phospholipids (Vick and Zimmerman, 1984). The products of membrane breakdown can also lead to further autocatalytic damage by the formation of harmful radicals or oxygen species, such as superoxide (Lynch and Thompson, 1984), singlet oxygen (Thompson et al., 1991 ) or lipid peroxy radicals. All three species are capable of inducing further membrane damage. Several enzymes, such as superoxide dismutase, ascorbate peroxidase, monodehydroascorbate reductase, catalase and peroxidase (Bowler et al., 1991; Bowler, Van Montagu and Inze, 1992), are involved in the detoxification of oxygen radicles and derivatives.
No investigations have been carried out on this topic in cassava. However, the rapid appearance of vascular streaking in harvested cassava roots has been suggested by Tanaka et a/. (1983) to be a result of membrane disorganization resulting in a loss of cellular compartmentation. One detailed study of lipid changes in harvested cassava roots has shown a progressive decline in phospholipid content indicating membrane degradation. It was speculated that these changes indicate structural alterations to membranes which might allow the interaction of substrates and enzymes producing dark streaking (Lalaguna and Agudo, 1989).
Overall, wound responses result in the restoration of the integrity of the damaged plant. The processes involved include; the formation and movement of signals from damaged tissues, the perception by undamaged cells of these signals, which results in the activation of pre-existing enzymes and in gene expression, and the formation of a wide range of enzymic and other proteins concerned with defence, containment and repair. Wounding also induces the biosynthesis of further signal substances which amplify and sustain the primary effects and lead to the coordinated response. Some responses to wounding occur close to the wound, others take place at a distance. Some are initiated within minutes, others take place hours or even days after the damage has been inflicted. The spatial and temporal control of the overall wound response is an important feature of its function as a defence system.
As the normal internal environment of the plant is reinstated, wound signal formation is suppressed and the wound response processes are reduced.
Physiological deterioration in cassava roots appears to share many of the common characteristics of plant wound responses. However, the sealing and healing aspects necessary for survival seem poorly expressed and are less localized in harvested cassava roots (Booth, 1976; Rickard and Coursey, 1981). This may be associated with the fact that cassava roots have no function in propagation, unlike other root crop storage organs (see Introduction). Normal responses to tissue injury result in wound repair, which reduces and finally eliminates the signals from damaged cells that elicit the cascade of wound reactions. In harvested cassava roots held under low humidity storage conditions (less than 80 percent RH) the cascade of wound responses is sustained and extends through the whole root, leading to physiological deterioration. It would thus appear that wound signal formation is maintained and the wound response processes are not reduced under these conditions.
Early studies reported a limited variation in the susceptibility of cassava cultivars to physiological deterioration. More recent studies have demonstrated that environmental growth conditions have a significant effect (see Chapter Three) on the crop's development. Differences in root wound-healing properties have not been investigated in cassava or its wild relatives. It is possible that an adequate root healing response has been lost from cassava as this characteristic was not a selection priority.