Controlled atmosphere storage technology (CAST)
By R.L. Semple
INTRODUCTION
The use of residual contact insecticides for the protection of stored commodities and structural treatments, and the use of fumigation as an emergency disinfestation technique are components in an overall pest control strategy. However the most primary form of control, is adequate sanitation and hygiene within the warehouse and including its immediate surroundings, in storage bins, conveyance machinery, processing machinery (i.e., rice mills) and transportation.
Interest in alternatives to chemical control has been rekindled due to problems related to their use;
- Resistance to registered and available grain protectants resulting in control failure.
- Lack of replacement insecticides that are relatively inexpensive and safe, which includes not only pest control operators but consumers and the enrironment.
- Unacceptable pesticide residues especially imposed by some importing countries that are not recognizing codex MRL's.
- High development costs and time required for chemical manufacturers to register grain protectants for commercial use.
- The requirement for achieving nil tolerance for live insects in export grain commodities (including raw cereals and milled products).
- Environmental and social imp/ications.
ENVIRONMENTAL MODIFICATION TECHNIQUES
Stored grain, associated insects and microorganisms respire using atmospheric oxygen to convert carbohydrates in the grain into carbon dioxide and water. In poorly ventilated bulk storage systems, the heat generated by this reaction is not easily dessipated due to the insular properties of the grain. Consequently, temperature rises in localized regions or ``hot spots", respiration rate increases and the rate of deterioration is accelerated.
This phenomenon can be best illustrated by the simple oxidation equation of glucose (starch of grain changes to glucose before most organisms digest the food) in air.
{kind=link}
If we review the ratios of atomic weights of the elements involved, we can compute the following relationship
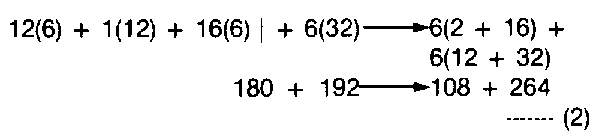{kind=link}
Where 180 parts of cereal plus 192 parts of oxygen, evolves 108 parts of water plus 264 parts of carbon dioxide. Any units of weight can be assigned to these values, so long as they remain constant. (atomic weights of carbon = 12; hydrogen = 1; oxygen = 16).
The breakdown of 180 grams of cereal produces 2820 kJ of free heat (1 KCal = 4.1868 kJ; 1 BTU = 1.05506 kJ), which must be dissipated to the surroundings, the zone of generation being at the higher temperature. Deteriorating grain creates hot spots that may be detected by temperature sensing thermocouples or resistance thermometers (thermistors).
The respiration of dry, uninfested grain is low, but in the presence of insects, the rate of respiration as measured by the production of carbon dioxide, significantly increases and is greater at elevated temperatures and moisture contents. Most storage fungi are unable to proliferate at relative humidities below 70%, and in such cases when insects are not present, the respiration rate is similar to that of dormant seeds.
Most animals and plants require oxygen for their respiration and will die or cease to grow at low oxygen tensions. Some organisms (such as yeasts and bacteria), are capable of anaerobic respiration, breaking down carbohydrates to substances such as lactic acid, acetic acid or alcohol, but producing less heat than aerobic respiration.
{kind=link}
In dry grain stored in air-tight conditions, an insect infestation will use up available oxygen and become asphyxiated (they are essentially obligate aerobes), if the oxygen concentration falls below 2% by volume of the intergranular air (Bailey, 1965). Fungi can still grow at oxygen concentrations down to approximately 0.2% (they are micro-aerophilic), while yeasts (anaerobic), can proliferate very rapidly at an oxygen tension between 0.5 -1.0% under suitable temperatures, causing fermentation.
Safe storage is therefore achieved by keeping the grain under conditions in which spoilage organisms are not able to rapidly develop or become established. Variables that can be manipulated and controlled are temperature, relative humidity and therefore grain moisture content (EMC), and the gaseous composition of the storage atmoshphere.
Table 1. Carbon dioxide concentrations as an index of infestation.
| CO2 CONCENTRATION % per 24 hrs | INDICATION OF INFESTATION |
| 0.25 | Uninfested grain with less than 15% m.c. |
| 0.30 | Clean grain |
| 0.35 - 0.50 | A slight insect infestation or a rater high infection of microorganisms |
| 0.50 - 0.90 | Close observation required |
| 1.0 | Limit of dangerous storage conditions |
| 1.0 | Highly unsuitable storage conditions |
DETERMINING GRAIN ACCEPTABILITY FOR STORAGE
Carbon dioxide evolution is directly related to dry matter loss (DML) of grain as a function of moisture. Teter (1979, 1981) stated equations for paddy as
log (CO2) = 0 44 Mw - 6.08 for Mw 13.3- 15.6%
log (CO2) = 0 21 Mw - 3.04 for Mw 10- 13.2%
log (CO2) = 2.33 sin | 7.5(M - 13) for Mw 15.7-25%
where Mw = % moisture wet basis.
The general equation is
log(CO2) = AMW- B (Hall,1970) -------(4)
where CO2 = mg CO2 per 100 gm dry matter day m = % moisture wetbasis
A & B = grain constants as shown in Table 2 (Appendix I).
Carbon dioxide evolution is a direct ratio of DML, such that for each gram CO2 evolved, 0.682 gram of dry matter is lost (180 C6H12O6 to 264 CO2; 180/264 = 0.682).
If follows that 1 kg of CO2 generation gives;
- 0.682 kg of dry matter loss
- 0.409 kg of H2O
- 10,680 kJ of heat generated (1000/264 x 2820 kJ)
The acceptability of grain for storage has been stated that the deterioration at the conclusion of storage should not exceed 0.8% DML (Teter, 1981), or in the U.S., 0.5% DML.
This level can be computed from
% DML per day = (CO2).10-3 (0.682)
1. EFFECTS OF MODIFIED ATMOSPHERES ON STORAGE INSECTS
Monro (1967) has summarized some of the original observations of Dendy and Elkington (1920) concerning the effects of air-tight (hermetic) storage on grain insects.
1. Grain insects in hermetic inclosures succumb as soon as the oxygen has been used up with a corresponding increase in the amount of CO2 being produced.
2. The gases present under normal sealed conditions are oxygen, nitrogen and carbon dioxide.
3. The amount of CO2 given off by live cereal grains in hermetic enclosures is directly related to moisture content and temperature, and above a critical point of moisture, the production of CO2 increases very rapidly.
4. Above a certain critical level of moisture content, grain stored under hermetic conditions creates an environment lethal to insects (and other aerobic organisms) quite rapidly, but at lower moistures, the rate of "self sterilization" takes a comparatively longer time to achieve.
5. The amount of oxygen consumed by live cereal grains of low moisture is greater than the amount of CO2 given off (i.e., a respiration quotient less than unity).
6. At approximately 30�C, 100 Sitophilus oryzae give off about 30 milligrams (i.e., nearly a fifth of their own body weight) of CO2 in 24 hours, while at lower temperatures (20-21 �C), the amount of CO2 evolved is reduced to about 9.4 mg in 24 hours.
7. Sitophilus granarius is a less active weevil as compared to S. oryzae, and on a weight for weight basis, the amount of CO2 evolved is less. The respiratory quotient (RQ or ratio of CO2 given off to the amount of O2 consumed) for S. oryzae is 0.773, while for S. granarius, it is around 0.815.
8. It is generally accepted that the absence or lack of oxygen is sufficiently lethal to weevils, without taking into account the presence of increased concentrations of CO2, although they may remain alive for extended periods under low oxygen tensions.
9. However, CO2 does exert a lethal effect on weevils irrespective of depleted oxygen tensions such that at approximately 30�C, S. oryzae was killed in less than 12 days in an atmosphere containing 14.0822.56% CO2, although around 14% 02 still remained.
10. Pure (moist) CO2 is less fatal than CO2 with a small admixture of O2, where pure CO2 acts almost instantaneously as a narcotic under the influence that Sitophilus spp. remain in a state of "suspended animation" without losing their power of recovery.
Sealed grain storage therefore has the following advantages:
- It sterilizes the grain, killing insects in all stages and other vermin that may be present.
- It prevents absolutely, the entry of insects and rodents and therefore obviates the problems associated with further applied control measures.
- It prevents grain of high moisture content from becoming mouldy and from heating, although it does not prevent anaerobic respiration and the associated rise in acidity.
Table 2A. Constants A and B for equation (4).
| Grain | Moisture Range | A | B |
| Maize, dent | 10.0 -13.2 | 0.17 | 2.00 |
| 13.3 - 17.0 | 0.27 | 3.33 | |
| Sorghum | 10.0 - 13.2 | 0.125 | 1.65 |
| 13.3 - 17.0 | 0.32 | 4.19 | |
| Rough rice | 10.0 -13.2 | 0.21 | 3.04 |
| 13.3 - 17.0 | 0.44 | 6.08 | |
| Brown rice | 10.0 - 13.7 | 0.17 | 2.67 |
| 13.8 - 17.0 | 0.44 | 6.41 | |
| Polished rice | 10.0 - 14.1 | 0.16 | 2.83 |
| 14.2 - 17.0 | 0.49 | 7.48 | |
| Wheat, soft | 10.0 - 14.0 | 0.09 | 1.35 |
| 14.1 - 17.0 | 0.36 | 5.14 |
- It prevents the absorption of moisture from the humid ambient atmosphere, so that if grain is initially stored in a dry condition, it remains dry.
The general conclusions are that grain stored in air-tight conditions can be maintained without appreciable deterioration in grain quality and germinative capacity, so long as it is initially dry. Grain above 17% m.c. deteriorates in time, while grain below 17% m.c., remains viable for 6 months or even a year, but shows low rates of germination and is therefore unsuitable for seeding and malting. At temperatures of 25�C, grain of 24% m.c. Ioses Its germinative capacity completely in 6 weeks, while grain stored at the same moisture content but at 15�C, loses its germinative capacity in a longer period (19 weeks).
The diffusion theory in insect respiration:
Wigglesworth (1966) stated that;
"Although the tracheae are permeable to oxygen throughout their length, the abundant supply of tracheal capillaries in and around the most active tissues leaves no doubt that most of the oxygen enters the organs through these endings but it is only comparatively recent that the diffusion theory for insect respiration in the conveyance of oxygen from the spiracles to the tracheoles has been widely accepted."
Essentially, the elimination of carbon dioxide is performed in a similar manner. Generally, the amount of CO2 produced by the insect is usually less than the amount of oxygen absorbed. The rates of diffusion is proportional to the square roots of their densities and since they do not differ dramatically, the theory of diffusion therefore accounts for both oxygen uptake and carbon dioxide elimination.
CO2 diffuses more rapidly (presumably) through insect cuticle of the body (especially when this is thin), and through the larger tracheal trunks, than oxygen entering via these routes, and it is generally safe to say that in most terrestrial insects, oxygen is taken in by the tracheae.
Tracheal respiration is compounded by the requirement for conserving body moisture, since most of the water loss in insects occurs by evaporation through the spiracles.
The function of the spiracles has been termed the "diffusion control of insect respiration". When the spiracles are closed, any active organ or tissue can draw on oxygen from any part of the system, and oxygen levels can be re-established by the opening of a single pair of spiracles. Normally, spiracles are kept closed and are only opened long enough for the insect to be supplied with oxygen. If an insect struggles or is under stress, or the rate of metabolism is increased due to elevated temperatures, or at the height of digestion and egg production in females, they are opened more frequently and remain open longer with other spiracles being activated, which results in increased rates of water loss.
Spiracular movement is regulated by specific ganglia of the ventral nerve cord and the brain, and are stimulated by CO2 such that an exposure to 2% CO2 incluces the spiracles to remain permanently open, as well as by the accumulation of acid metabolites due to oxygen lack. In normal respiration, the reduced tensions of oxygen and accumulation of CO2 (except in thin-skinned insects where most of the CO2 diffuses through the general surface of the body, and consequently cannot serve as a respiratory stimulus) cooperate in the control of respiration.
Gaseous diffusion is adequate for small insects, but is insufficient for actively running or flying species with a high rate of metabolism and requirement for energy. A certain degree of mechanical ventilation is added to compensate for the higher requirement for oxygen.
The main function of collapsible tracheae and airsacs (tracheae with large dilatations which occupy a great part of the body cavity as in many Orthoptera and Hymenoptera) is the ventilation of the respiratory system. They are ventlated by the respiratory movements of the rigid body wall that encloses them, and to some extent by general body movements, such as during flight. Therefore, the greater part of the tracheal system is maintained full with a gas similar in gaseous composition to that surrounding the insect, but the supply of oxygen to the tissues is still effected by diffusion along the tracheal branches extending from these airsacks.
The extent to which the tracheal system is ventillated varies dramatically between different insect species, or in the same insect in different physiological states. Respiratory movements of insects are brought about by special musculature generally confined to the abdomen, but in some insects (aquatic), ventillation is maintained by aspirating movements of the metathorax.
So it can be seen that when oxygen requirements are great, ventilatory pumping movements are initiated and the rythm of opening and closing of spiracles is modified in such a way that a directed stream of air is forced through the system, a system which necessitates a complex nervous co-ordination between pumping movements and regulation of spiracular movement.
Either a lack of oxygen or an excess of carbon dioxide may stimulate the respiratory centres and cause hyperpnoea (= abnormally rapid or deep breathing).
The secondary respiratory centre located in one of the thoracic segments has an overriding action and control movements in the entire insect, can be stimulated by CO2 tensions of 0.2 - 3.6%, while the primary centres (usually found in the segmental ganglia and control the movements of their own segments, i.e., isolated segments of the abdomen can perform respiratory movements) can be activated at 12-15% CO2, while ventillation in other insects can occur at CO2 tensions of 10%, even when the insect is at rest. It is highly probable that a chemical stimuli is acting in all cases, and is probably an increased acidity in the nerve centres, due either to excess CO2, or an accumulation of acid metabolites due to oxygen lack.
Bailey (1965), and Bailey and Banks (1974), demonstrated that the oxygen tension should be maintained below 4.5% O2 to produce a lethal atmosphere for even the most susceptible stored product insect, however, a limit of approximately 2% O2 has been predicted to ensure mortality of stored products insects that have been so far tested (Oxley and Wichenden, 1963; Banks, 1979).
Girish (1978) has made several observations on the susceptibility of various life stages of Trogoderma granarium Everts under varying exposures of gastightness at optimum conditions for development (35�C; 11.5% m.c.). These observations are summarized below.
1. Susceptibility to air-tight conditions varied from stage to stage and age of larvae.
2. No life stage was capable of surviving or completing development to the next stage under airtight conditions and 100% filling ratio of the container, except for pupae.
3. Reduction of free air space or headspace decreases available oxygen and is depleted more quickly than if insects are maintained in an empty container.
4. Susceptibility to low oxygen tensions produced biologically by grain and insect respiration was egg < first instar larva < adult < full grown larvae and pupae.
5. Oxygen consumption was greater than CO2 production for all stages.
6. Carbon dioxide production and O2 consumption is suppressed by the depleted O2 and accumulated CO2 under airtight conditions.
7. Grain respiration is much less than different life stages under air-tight storage conditions.
8. Under air-tight conditions, the levels of CO2 and O2 to achieve complete mortality was:
| Life Stage of T. granarium | : Gas Concentrations (%) |
|
| : CO2 | : O2 | |
| Adults | 12.56 | 3.39 |
| Full-grown larvae | 14.62 | 1.06 |
| First stage larvae | 12.34 | 5.35 |
| Eggs | 2.49 | 16.77 |
Observations (3) and (5) from this study require further clarification. Referring back to the oxidation equation of glucose (hexose polysaccharide) the volume of CO2 produced is equal to the volume of O2 consumed in the respiratory process, which represents a respiratory quotient (RQ) of 1 or unity.
This is rarely obtained in air tight conditions where the filling ratio is 100% (RQ = 0.7) mainly because CO2 is absorbed by the grain (in greater amounts than absorbed oxygen relative to the net volume of grain), and is therefore not measured in the interstitial airspace. Once the filling ratio in airtight containers is reduced (to < 5%), the RQ approaches unity (see Table 3). For this reason, even with high population densities of insects (assuming anaerobic respiration is not evident) in completely sealed containers, oxygen depletion to a lethal 2% is accompanied by a rise in CO2 to rarely more than 15% (RQ of approximately 0.75).
Table 2. Influence of moisture content and temperature on the respiration and mortality of S. oryzae in wheat (in relation to equilibrioum relative humidity) (Results from Lindgren, 1935)
| Wheat moisture content ( % ) | Respiration of uninfested wheat (mg CO2/48 hours/100 g of wheat) | Respiration of S. oryzae (mg CO2/48 hours/1 g of weevils) | ||
| 25�C | 35�C | 25�C | 35�C | |
| 8.7 | trace | trace | 151.6 | 129.0 |
| 10.7 | trace | 0.44 | 300.0 | 215.4 |
| 14.0 | 1.34 | 2.90 | 412.8 | 522.6 |
| 15.2 | 2.00 | 4.48 | 403.8 | 578.8 |
| 17.4 | 26.4 | 42.4 | 409.2 | 464.0 |
Table 3. Apparent Respiratory Quotient (ratio of evolved CO2 to oxygen consumed) of equal numbers of S. oryzae adults in jars of equal size but containing varying proportions of wheat, i.e., the filling ratio.
| Proportion of container occupied by wheat ( % ) | Apparent respiratory quotinet |
| 100 | 0.69 |
| 75 | 0.78 |
| 50 | 0.86 |
| 25 | 0.94 |
| 5 | 1.01 |
Extracted from "The effect of restricted air supply on some insects which infest grain" by T. A. Oxley and G. Wickenden Ann. Appl. Biol. (1963): 51, 313-324.
The RQ of T. granarium approaches but is less than unity, as shown in (5). Girish (1978) and Bailey (1955) suggest it would be advantageous to completely fill air-tight silos because of more rapid O2 depletion (lack of air space means an interstitial air space of 40% of total volume with wheat), but Oxley and Wickenden (1963) found very little difference between filling ratio and rate of O2 depletion.
In air tight or hermetic storage systems, it is therefore oxygen depletion by the respiration of insects themselves that is the lethal factor rather than the accumulation of CO2.
With externally generated controlled atmospheres, the combinations of gaseous mixtures is easily manipulated and may produce synergistic effects. Atmospheres containing 15% O2/36% CO2/balance air have been found to be lethal to Tribolium castaneum (Herbs") and Plodia interpunctella (Hub.) larvae after an exposure of 10 days (Herein and Press, 1968). Calderon and Navarro (1979) exposed T. castaneum adults for 24-120 hrs at 57% RH and temperatures of 26�C and 30�C, atmospheres containing O2 concentrations of 2 - 5% with 5 - 35% CO2. A clear synergistic effect was found, since mortality at given O2 concentrations was higher when the level of CO2 was increased. Mortality was also subsequently higher at the higher temperature and if the exposure period was longer. With a 5% O2 concentration, no mortality was observed, but the addition of 35% CO2 caused 100% mortality at both temperatures. To obtain an LD95 in atmospheres containing 2% O2, the addition of 10% CO2 is required, but 5% O2 plus 15% CO2 produced only negligible mortality. When CO2 is present, O2 concentrations are depleted more quickly and adults die at higher concentrations. Both O2 depletion and presence of CO2 increases water loss which increases mortality (Navarro and Calderon, 1974). Enhanced insect mortality with reduced O2/increased CO2 concentrations in the atmosphere emphasizes the advantage of using CO2 to obtain low O2 tensions in preference to purging with N2.
Calderon and Navarro (1980) further substantiated the synergistic effects of the addition of CO2 to low O2 atmospheres on adult mortality of T. castaneum and Rhyzopertha dominica (Fab.) which appeared more susceptible (100% mortality at 2% O2 with the addition of only 10% CO2).
Results on the effects of CO2 to reduced O2 atmospheres on 0-24 hr eggs showed that with T. castaneum, 100% mortality was achieved at 2 and 4% O2 without CO2 (96 hr exposure) while even 8% O2 inhibited almost 50% of the eggs to hatch (it should be noted that no adult mortality was achieved in O2 alone above 4%).
The addition of CO2 to given O2 atmospheres had little effect on egg mortality, but increasing CO2 concentrations (from 5-30%) did display an additive effect in the range of O2 concentrations tested (2-8%), but failed to exhibit any synergistic effect on egghatch. Presumably greater water loss in adults in high CO2 atmospheres as compared to eggs is the reason for differing response. Eggs are significantly less effected by low relative humidities than adults of Ephestia cautella and T. castaneum.
These studies have established the toxicity of various gaseous combinations beyond those established in earlier publications by Bailey. O2 concentrations of 3-4% or between 40-60% in the presence of atmospheric O2 concentrations (21 %) at 23-32�C with 14 day exposures depending on the species.
Various workers have found that mixtures of 90/10 (CO2/O2) were as lethal as 98/2% mixtures against T. confusum, and 80/20% mixtures were much more toxic than 100% CO2 against S. oryzae and S. granarius adults, both tests performed at 26.7�C. (All Niazee, 1971; Lindgren and Vincent, 1970).
Jay (1983) found that 60% CO2 in air is more effective against the internal feeders or primary grain pests such as Sitophilus spp., R. dominica and T. granarium, while atmospheres containing 100% CO2 is more effective against the external feeders or secondary grain pests, such as T. castaneum, Oryzaephilus surinamensis, Trogoderma variable and T. glabrum, presumably because of adsorption and desorption of CO2 on grain. The practical inference is that different gaseous mixtures could be used depending on species composition present in storage.
Bell (1983) has studied the toxicity of O2 in high concentrations of CO2 on the eggs of Ephestia cautella and E. Kuhniella at 15 and 25�C.
At 25�C, gas mixtures containing more than 40% CO2 killed all eggs (aged 2, 20 and 44 furs) within 48 hours. Egg immediately after diposition are more susceptible to atmospheres containing 40% CO2 either in the presence or absence of O2, while the susceptibility to this level decreases as embryogenesis proceeds, in the presence or absence of O2. The presence of O2 plays some part in the toxic action of CO2, since susceptibility is prolonged in the presence of 4-20% O2, (up to 44 hrs), while in the absence of O2, susceptibility to concentrations of 40% CO2 and above decreases in 20 hour old eggs.
At lower temperatures, longer exposures are required to obtain complete mortality with CO2 in the presence of O2 than in its absence.
The dessicating action of CO2 can be ignored for eggs since its action affects spiracular control but if development proceeds during exposure, mortality will be achieved when a more susceptible stage is reached. CO2 is actively toxic at higher concentrations (unlike N2, it does not rely on anoxia to be lethal) and may be more toxic in the presence of oxygen. The speed of action of CO2 in air is only slightly faster than CO2 alone, but is enhanced substantially at lower temperatures, where purging storages with CO2/O2 mixtures may achieve better results than purging with CO2 alone. Also, purging with CO2 remains effective for longer than N2, since the atmosphere is diluted progressively as air leaks back into the purged structure.
Jay (1983) claims storage insects are more susceptible under field applications than in laboratory evaluations, because of the gradual decline in concentrations during the ventillation phase. At 70% RH, 77% CO2, the exposures required for T. granarium and pupae of s. granarius is one month, but because of natural gas loss, an indefinite exposure for T. granarium maybe warranted. However, he emphasized the dearth of knowledge of CAST as related to grain moisture and relative humidity, especially conditions operative in the humid tropics.
Insects infesting stored products are capable of withstanding extremely low humidities, an important aspect of their structure and physilogy which allows rapid changes of O2 and CO2 while restricting water loss by efficient respiratory systems. Navarro (1978) suggests that with respect to controlled atmospheres, the structure and physiological characteristics of each species should be taken into consideration. For example, Ephestia cautella pupae in 3% O2 with low RH, mortality is due to dessication through effects on the spiracular regulatory system, but if the RH remains high, some survival is possible (Navarro, 1978).
At lower oxygen tensions (1 % O2 for E. cautella or 0.12% for T. castaneum adults) insect mortality occureed at high RH, but when the RH was low, the desiccation rate increased causing mortality in a shorter period, such that a relationship of higher RH, lower O2 to achieve similar levels of mortality exists.
Navarro and Calderon (1980) have emphasized the requirement of investigating temperature and relative humidity requirements for more efficient use of CAST. At a given atmospheric gas composition, the higher the temperature, the shorter the exposure time reguired to achieve an LD95, and is very pronounced in the range from 15-21�C. Since different temperature gradients exist in grain bulks, the lowest recorded temperature should govern the length of exposure period required. Also since the response of storage insects differs to CAST, the most tolerant species present will influence the system used.
Controlled atmosphere treatments appear more effective when the moisture content of the grain is low. Due to moisture migration, moisture contents in localized parts of the bulk maybe considerably higher than the remaining bulk, constituting the need for applied gas concentrations to achieve effective control of insects residing in higher moisture grain.
Damp pockets of grain can cause insect survival with CAST, and although Navarro and Calderon (1980) claim drying of the grain bulk for obtaining efficient insect control with modified atmospheres is impractical, the requirement for grain drying in the humid tropics is a prerequisite for good storage.
The effectiveness of sealing techniques to prevent excessive use of gas mixtures has been established, but to render storages completely gaslight is an expensive if not impractical undertaking.
{kind=link}
If aiming for a lethal modified atmosphere of less than 2% O2 under hermetic storage, and if the walls of the container allow diffusion of oxygen From the atmosphere, Insects will also be killed dependent on the rate of diffusion (which is also dependent on the difference between internal and external concentrations) and the initial population density (Oxley and Wickenden, 1963).
There is also the possibility of surviving insects aggregating around leaks at higher O2 tensions (Navarro, et a/., 1979). In experiments where O2 levels were measured at 17% around leaks, O2 concentrations approximately 90 cm from the leak dropped to a lethal 1.6% (Navarro, et. al., 1976).
Distributions of R. dominica, S. oryzae and 0. surinamensis have been monitored under a range of leakage rates and gas gradients in a simulated laboratory silo (Navarro, 1977). Results using atmospheric gases suggest that none of the insects examined are able to migrate to high oxygen concentrations (ca. 21 %) in a nitrogen or carbon dioxide enriched atmosphere. Insect movement is apparently inhibited in very low oxygen concentrations ( c 1 %) in nitrogen, and by high CO2 enriched atmosphere (>65%) in air with O. surinamensis (Navarro, et. al., 1981).
Relative short purge periods with N2 to achieve 1 % O2 (approximately 12 hrs in a 2000 tonne bin of wheat at a purge rate of 3 m3.min-1 (Banks and Annis, 1977) dispersion from low to high O2 tensions existing around cracks in the bin structure is remote, but insects already present in this region may survive the treatment (Navarro, et al., 1981).
The response of O. surinamensis to CO2 atmospheres, dispersal is restricted to concentrations up to and including 66% CO2. Other studies with Tribolium spp. have shown attraction to CO2 concentrations ranging from 0.25- 15%, while repellency occurs between 60 - 90% CO2.
In practice, the purging phase to achieve 20% CO2 is short enough ( < 24 furs) to severely restrict insect dispersion to regions of low CO2 concentration.
These observations suggest there is an advantage in using CAST for insect control, since the infestation will not redistribute itself to regions of high O2 or low CO2 concentrations in the period of time required to obtain the controlled atmosphere.
A potential problem of CAST is the development of resistance, since stored grain insects do possess the genetic potential to develop CO2 resistance (Bond and Buckland, 1979), although no field strains have exhibited this ability. Insects are essentially obligate aerobes, which may indicate a potential advantage of using N2 to create lethal O2 tensions, in preference to the toxic action of CO2.
2. EFFECTS OF MODIFIED ATMOSPHERES ON MICROORGANISMS
Christensen (1978) listed the advantages of CAST as inhibition of aerobic fungi, prevention of mycotoxin production and conservation of desirable quality factors of the grain, while some disadvantages are the elimination of microorganisms that compete under aerobic conditions, development of populations of certain aerobic microorganisms before the atmosphere has been sufficiently modified, expensive and complex technical facilities and capabilities, and generation of some adverse quality factors.
Busta, et al., 1980 consider the following as major considerations influencing the effectiveness of CAST such as the types and numbers of microorganisms present including both storage and field fungi, yeasts, including Candida and Hansenula spp, and bacteria including the genera Lactobacillus, Clostridium, Bacillus and general groups such as the coliforms.
Other important considerations are grain type, and primary physical parameters, both intrinsic and extrinsic factors, influencing microbial activity such as grain moisture or water activity, storage temperature, atmosphere surrounding the grain during storage and storage duration.
Temperature and moisture requirements of fungi have been sufficiently reviewed elsewhere (Christensen and Kaufmann, 1974; Christensen an Kaufmann, 1977, Northolt, 1979).
In commodity modified or biologically produced atmospoheres, respiration and basic metabolism reduce available O2 and increase CO2, where a rapid increase in the production of CO2 above a grain moisture content of 15% mc was accompanied by a relative increase in the number of moulds on the grain (Milner, et al., 1947). If fungicidal or fungistatic substances are applied, respiration remains low even at 20% mc.
Similar reductions can be achieved with CAST by the precise and constant maintenance of CO2, N2 or O2 at specific pressures or under a partial vacuum, but is expensive and technically demanding, while the atmosphere developed under air-tight storage is self developed and dynamic, depending only on the activities of biological organisms and the grain itself (Busta, et at., 1980). Aflatoxin inhibition has been demonstrated in atmospheres containing 90% CO2/10% O2 at 28�C (Shih and Marth, 1973).
Table 5. Relationship of moisture content to respiratory rate and mould growth (Regent wheat after respiratory trials at 30�C). From Milner et al., 1947.
| Initial Moisture Content (%) | Respiratory rate, (mg CO2/100g grain) | Mould colonies (per mf of grain) |
| 12.3 | 0.07 | 0.5 |
| 13.6 | 0.11 | 0.1 |
| 13.8 | 0.23 | 0.1 |
| 1 4.5 | 0.57 | 0.4 |
| 15.4 | 2.53 | 4.8 |
| 16.3 | 23.35 | 396 |
| 16.8 | 20.3 | 209 |
| 18.5 | 111.0 | 2,275 |
| 20.8 | 604.9 | 11,300 |
| 25.2 | 1,724.8 | 37,500 |
| 30.5 | 1,282.0 | 63,500 |
| 38.6 | 4,666.5 | 67,000 |
Table 6. CAST inhibition of growth or toxin production on corn and wheat *
| Grain Type | % Composition of Atmosphere O2/CO2/N2 |
|
| Corn | 0.5/13.5/84.81 | (at 27�C) |
| 0.3/0/99.7� | (at 27�C) | |
| Wheat | 0/0/1003 | (22 - 32�C) |
| 4.5/0/95.53 | (22 - 32�C) | |
- Wilson and Jay, 1975.
- Wilson, Huang and Jay, 1975.
- Shejbal, 1979
- Extracted from Busta et al., 1980.
Various interactions between temperature, relative humidity, and gaseous composition have been evaluated and it appears that a 3-way interaction exists. The required CAST system and gas composition will be very dependent on temperature and relative humidity (or water activity, Aw) operating in any particular storage situation.
Lillehoj et al, (1972) have estimated mean germination of Penecillium martensli in various gas mixtures after 16 hours. It appears normally distributed about 30�C where mean percent germination was approximately 37% in air, 30% in 20% CO2, 20% O2 and 60% N2., approximately 10% in 40% CO2, 20% O2, and 40% N2., and approximately 2% in an atmosphere containing 60% CO2, 20% O2 and 20% N2
3. EXTERNALLY MODIFIED CONTROLLED ATMOSPHERE STORAGE
The techniques involved in modified or controlled atmosphere storage of grain involves the alteration of the normal atmospheric gases present (21% O2, 0.03% CO2, 1% argon and other gases, balance nitrogen) so as to obtain an artificial atmosphere that is insecticidal and prevents mould growth and quality deterioration of the commodity.
Three types of controlled atmospheres have been under investigation by the Commonwealth Scientific and Industrial Research Organization (CSIRO), Canberra, Australia which are:
- Low oxygen atmospheres generated by purging with nitrogen, and maintained by addition of further nitrogen to compensate for air ingress (less than 2% O2).
- Low oxygen produced and maintained by the combustion of hydrocarbon fuels (13% CO2, 1 % O2, balance N2)
- High carbon dioxide atmospheres (35-80%) either allowed to decay through its active range or if this is too rapid, maintained with further CO2.
The methods used are based on the ancient process of hermetic storage where the respiratory activities of insects and moulds alter the atmospheric composition in a gas-tight structure by gradually using up the available oxygen and depleting it to around 2% with a subsequent rise in CO2 to around 13%, (anaerobic fermentation in damp grain under hermetic conditions may increase CO2 as high as 60%). It should be emphasized that the technology involved in CAST differ substantially from that of modified atmospheres generated by biological processes within a sealed structure, such as hermetic storage (see separate section on airtight storage), whereby the appropriate atmospher is supplied externally and introduced into the structure, and additional gas added if necessary to maintain the lethal atmosphere.
The first successfully recorded large scale treatments in Australia occurred during the World War I, but the process was not used widely again until 1979 (Banks, et al., 1980).
Externally generated atmospheres for disinfesting stored bagged grain in Australia from 1917-1919 was probably the first extensive use of such a system in the world (Winterbottom, 1922). Disruption of world trade had resulted in large stocks being stored in Australia where approximately 60,000 tonnes was treated with low oxygen atmospheres genearated from burning coke in a modified producer gas generator.
3.1 Modified atmospheres using liquid nitrogen: (From Banks and Annis, 1977)
Controlled atmosphere storage with nitrogen can be viewed in two phases, the purge phase, where the desired atmosphere is obtained in the bin or structure, and the maintenance phase, where this atmosphere is maintained for the desired exposure.
Liquid nitrogen supplied under pressure by road tanker is passed through a heat exchanging facility where it is vaporized and bought to ambient temperature (within 2�C). It is then passed through a flowmeter (rotameter) and then into the gas introduction system or manifold. Gas flows of 3 m�. min1 has been suitable for purging bins from 300-7000 tonne capacity. Increasing the gas flow rate maybe less efficient in reducing oxygen tensions during the purge phase in silos other than those fitted with aeration ducting, since pockets of air may remain with high O2 levels before being removed by diffusion and convection. thus wasting gas. A vent of at least 50 cm� must be left open in the roof during the purge in order to prevent dangerous pressure build up. A pressure release safety valve must also be installed to prevent any posit,ve pressures from endangering the structure.
Gas input must continue uninterrupted until the headspace has been reduced to 1 % O2. Purging in the grain mass occures as a defined front through the grain and direct displacement of the interstitial air. In the headspace, where free gas mixing occurs, decay is exponential.
The quantity of nitrogen required during the purge phase is dependent on the filling ratio and the porosity due to different commodities (i.e., barley requires higher gas input then wheat).
Once the headspace has been reduced to 1 % O2, the purge phase is discontinued, the top vents and introduction ports closed, and the maintenance input of gas commenced by addition of nitrogen in the headspace.
Maintenance gas is also supplied by liquid nitrogen in an insulated vessel on site and charged into the structure through an atmospheric heat exchanger. A simple continuous gas input system has been found superior to using a demand system which maintains a small positive pressure of 120 Pa in the storage, since the former used less nitorgen to maintain the same O2 tension.
Once the exposure period has been met, the atmosphere of a partly sealed structure will slowly return to normal atmospheric composition containing 21% O2, since air excluded by the maintenance phase leaks in.
This is dependent in levels of gastightness and weather conditions and may be accelerated by forced draught ventillation using fitted aeration equipment or simply providing screened openings (60 mesh) to restrict insect movement at the base and in the headspace. It should be emphasized that oxygen tensions should always be evaluated before entering storges that have been disinfested in this manner.
Choice of exposure period:
If disinfestation of grain is only required, prolonged CAST appears unnecessary. However, a perfectly sealed storage should be insect proof and therefore prevent reinfestation, but is dependent on the gastightness requiements with relation to the leak size (either one large leak or comprised of many smaller leaks) which may permit insect entry, and also their ability to locate suitable entry points.
Nitrogen storage reduces the formation of aflatoxins from the Aspergillus flavus oryzae group of storage moulds, and reduces total mould count. This may allow the safe storage of grains beyond the 12% mc limit that is imposed in Australia (where mould infection is not a serious problem compared to the humid tropics), and also reduces the hazards of heating in oil seeds as well as eliminating the risk of fire and restricting mould growth, but the little effect on the rise of FFA's. Shejbal (1979) has made the observation that the viability of malting barley could be preserved for long storage periods under nitrogen.
Table 7. Exposure periods and concentration requirements for modified atmosphere disinfestation of grain ( 12% me).
| Requirements | Atmosphere Source |
|
| Carbon Dioxide | Nitrogen | |
| Initial target concentration in storage | 70% CO2 in air (80%) | 1 % O2 in N2 |
| Final target concentration in storage | 35% CO2 in air (40%) | 1 % O2 in N2 |
| Period of exposure within these limits. | 25�C: 10 days | 24 wks at 15�C |
| 18 - 25�C: 14 days | 6 wks at 20�C | |
| 3 wks at 26�C | ||
| 2 wks at 30�C | ||
| 1 wk at 35�C | ||
SOURCE:
Banks and Annis (1977)
Banks et al., (1980)
* The exposure periods for nitrogen CAST are tentative, and based on S oryzae, the most tolerant grain pest to low oxygen atmospheres. If Tribolium spp or Oryzaephilus spp are present, shorter exposure periods maybe used, but if there is mixed populations, or any doubt to the indentity of the grain pests present, the recommendations for S. oryzae should be adhered to.