3.3.4 Mise en �vidence de spores dans des excr�ments de Prostephanus truncatus infest�s
Table
des mati�res - ![]() Pr�c�dente - Suivante
Pr�c�dente - Suivante![]()
Un insecte peut contracter une infection de protozoaires par ingestion (LIPA, 1974: WEISER, 1961). Eu �gard 4 ce mode d'infection, ;1 est important de savoir de quelle mani�re l'agent pathog�ne va se propager d'un h�te � l'autre au sein d'une population. S'agissant de P. truncatus , il existe deux possibilit�s de diss�mination des spores dans le substrat nutritif. Il y a d'une part, en un certain point, diss�mination dans le substrat d'une quantit� de spores relativement importante, cons�cutive � la d�composition d'un animal mort et infeste. Ces spores vont �tre ensuite propag�es peu � peu par ceux de ses cong�n�res qui se seront contamines en les ing�rant Les spores peuvent d'autre part p�n�trer dans le substrat nutritif par les excr�ments des animaux et �tre ensuite propag�es. Afin d'�lucider la question de l'introduction des spores dans les excr�ments de P. truncatus infectes, on a proc�d� � l'essai suivant.
Ayant pr�lev� dans l'�levage infest� 25 adultes de P. truncatus, on les a soigneusement nettoy�s afin de les d�barrasser des particules de farine. Cette pr�caution avait pour but d'�ter les �ventuels spores de protozoaires adh�rant au corps des insectes. Les col�opt�res ont �t� ensuite places s�par�ment dans des bo�tes de Petri en verre, dont le fond �tait recouvert d'un morceau de papier, et gard�s dans ces bo�tes jusqu'� leur mort. Apres leur mort, les sujets ont �t� examin�s en vue de d�tecter une �ventuelle infestation de protozoaires. Leurs excr�ments ont ensuite �t� retires des boites de verre et examines, sous forme de pr�paration, afin de d�celer 13 encore une infestation �ventuelle par des spores de protozoaires.
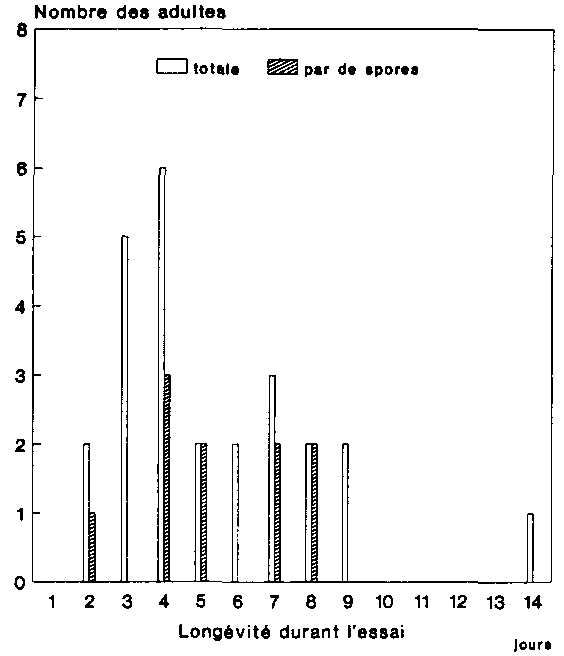
La figure 10 montre les spores mises en �vidence dans les excr�ments des 25 adultes de P. truncatus Des spores ont �t� d�couvertes chez 10 des anobies, lesquelles �taient toutes victimes d'une forte infestation de protozoaires. Il s'agissait en l'occurrence, et sans exception, de spores de Mattesia bien que quelques-uns des animaux examin�s aient �t� �galement atteints d'une infection mixte, provoqu�e par des Nosema sp. et des Mattesia sp. Chez 2 individus seulement, on a pu isoler dans les mati�res f�cales un nombre relativement �lev� de spores. Chez les 8 autres sujets, en revanche, on a d�nombr� moins de 10 spores de Mattesia seulement par pr�paration.
La figure 10 renseigne par ailleurs sur la long�vit� des sujets plac�s en observation. Si l'un deux a surv�cu durant 14 jours, la plupart �taient d�j� morts au bout de � ou 4 jours. Les excr�ments des P. truncatus adultes ayant surv�cu plus longtemps, � savoir entre g et 14 jours, ne recelaient pas de spores. Il convient de pr�ciser au sujet de ces r�sultats que l'infection de protozoaires, d�j� fort avanc�e, dont les animaux h�tes adultes �taient victimes, a tr�s certainement entra�n� des alt�rations physiologiques qui ont provoque � leur tour des troubles digestifs. Les animaux s�lectionn�s n'ont par ailleurs re�u aucune nourriture durant l'essai.
En r�sum�, on peut conclure que la propagation des spores par l'interm�diaire des mati�res f�cales de P. truncatus infest�s ne rev�t une certaine signification que dans le cas de Mattesia sp. Eu �gard aux faibles quantit�s de spores, d�couvertes en outre dans quelques parties d'excr�ments seulement, il est toutefois tris probable que les protozoaires se transmettent essentiellement par le biais de cadavres infest�s en d�composition et que la transmission par les mati�res f�cales demeure un ph�nom�ne tr�s limit�.
En compl�ment des observations individuelles pr�c�demment d�crites, on a pratiqu� deux essais d'inoculation destin�s � �tudier la dynamique de population de l'anobie P. truncatus Pour le premier de ces essais, on a utilise des col�opt�res infestes naturellement, et cela afin d'�tablir dans quelle mesure il �tait possible d'intensifier l'infestation naturelle de la population de ravageurs par des protozoaires en proc�dant � un traitement aux spores. vu que l'on trouve effectivement dans les greniers � ma�s des populations de ravageurs infest�es naturellement, cet aspect de la question rev�t une importance certaine du point de vue des conditions pratiques de la mise en oeuvre des protozoaires. Dans le cadre d'un second essai, on utilise des anobies exemptes d'infestation. La question primordiale consistait en l'occurrence � d�terminer si les protozoaires Etaient capables de s'�tablir dans la population de l'h�te � la suite du traitement aux spores, et, dans l'affirmative, avec quels r�sultats.
3.3.5.1 Inoculation pratiqu�e sur des Prostephanus truncatus infest�s naturellement
Pour observer la dynamique de population de P. truncatus infestes naturellement, on a pris � conteneurs, dans chacun desquels on a appliqu� � 60 col�opt�res, sans distinction de sexe, la pr�paration mixte aux spores. Parall�lement � cette exp�rience, on a pr�par� une variante non trait�e Les populations ont �t� alors plac�es en observations les imagos et les larves mortes �tant retir�s des conteneurs au bout de 4, 8, 12 et 16 semaines. On a ensuite �tabli sur 50 individus, repr�sentant chacun des stades, le pourcentage d'infection par les protozoaires.
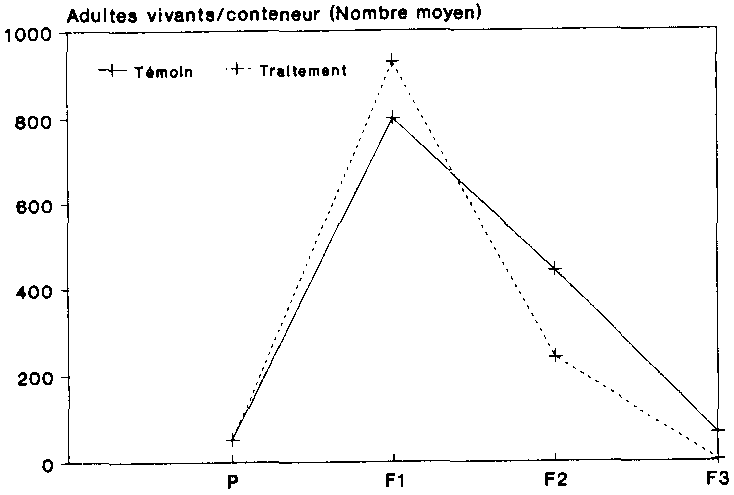
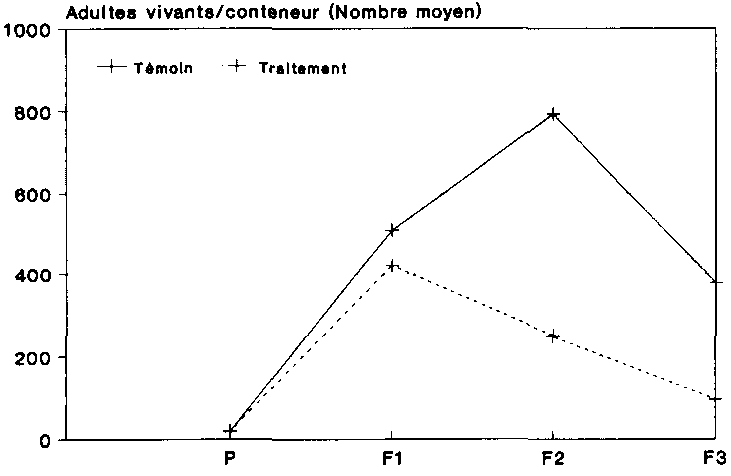
On trouvera sur la figure 11 une comparaison du nombre moyen d'adultes vivants pour chacune des variantes. Sous l'influence du traitement aux spores, la g�n�ration P a donn� naissance � une premi�re g�n�ration filiale de 929 adultes vivants, pendant que la g�n�ration P du t�moin produisait BOC col�opt�res. Dans la g�n�ration filiale F2, les populations trait�es comptaient 240 animaux seulement, alors que le nombre de sujets �tait presque deux fois plus �lev� dans les r�cipients non traites Sous l'influence des spores, on ne comptait plus en moyenne dans la g�n�ration F3 que � imagos vivants, alors que l'on d�nombrait 65 col�opt�res chez les populations non trait�es. Les populations-t�moins pr�sentaient par cons�quent une dynamique un peu plus stables que les populations trait�es.
Le recul accentu� au nombre d'adultes enregistr� au sein de F2 et de F3 s'explique par le fait que les imag�s aptes a la reproduction ont �t� s�par�s des populations assez rapidement apr�s maturit�. Selon SHIRES (1979), la dur�e moyenne du cycle �volutif de P. truncatus, depuis la larve jusqu'� l'animal adulte, est d'env. 28 jours dans les conditions de l'essai r�alis� ici (30 �C et 80 % d'humidit� relative).
Cette p�riode correspond toutefois exactement aux dates de la dynamique auxquelles on s'est conform� dans le cadre de l'essai. La plupart des adultes n'avaient donc que peu de temps pour la copulation et la ponte, ce qui fait que le nombre d'adultes a diminue de plus en plus nettement d'une date � l'autre.
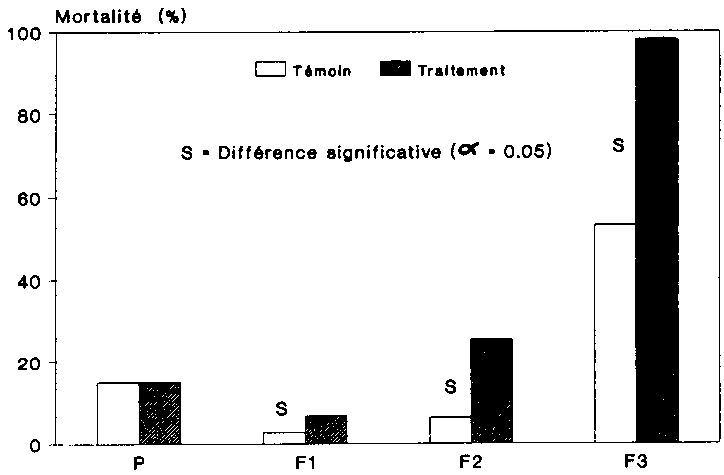
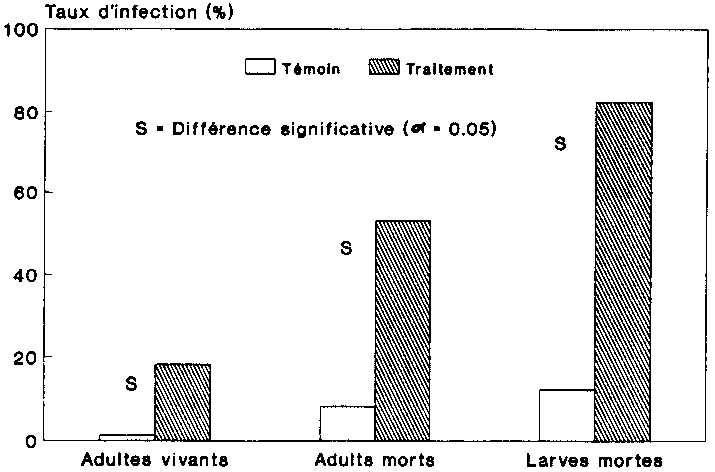
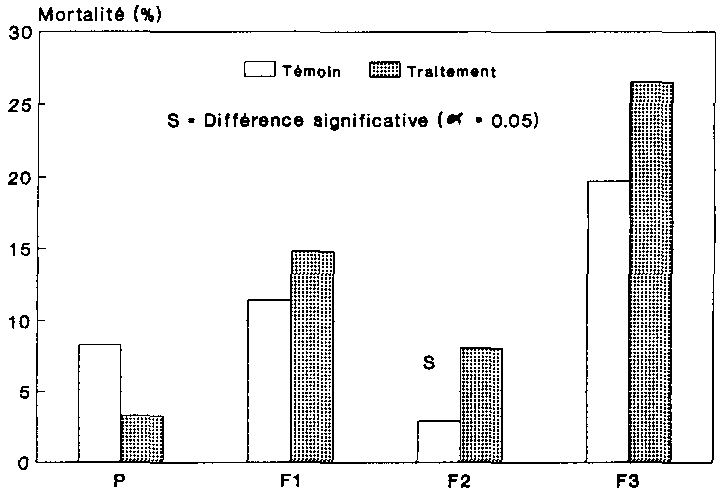
La figure 12 indique la mortalit� des adultes pour chaque variante. Il s'agit ici du taux de mortalit� moyen au sein des g�n�rations. La comparaison statistique du taux de mortalit� des deux variantes a �t� effectu�e au moyen du test Chi2. Les diff�rence significative sont indiqu�es sur la figure 12. Si, de F1 � F3, la mortalit� a augmente aussi bien au sein des populations non trait�es que des populations trait�es, le taux de mortalit� des adultes infestes par les spores (tait cependant beaucoup plus �lev�. Dans la troisi�me g�n�ration filiale, presque tous les col�opt�res traites, soit 98 %, �taient morts, alors que ce chiffre n'�tait que de 53 % d'adultes pour la population-t�moin.
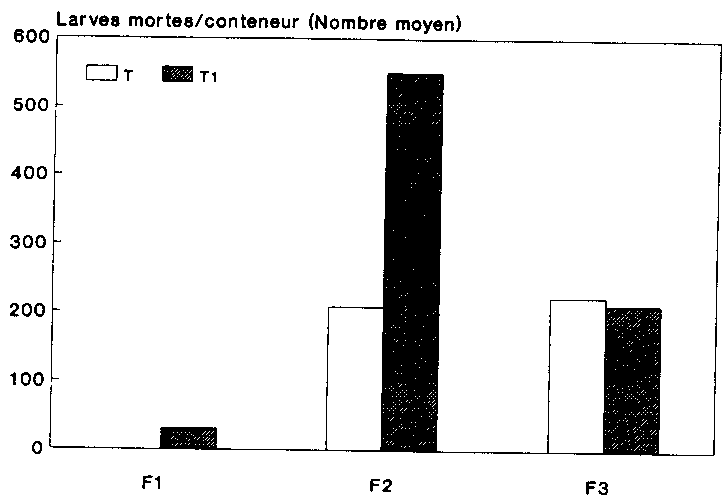
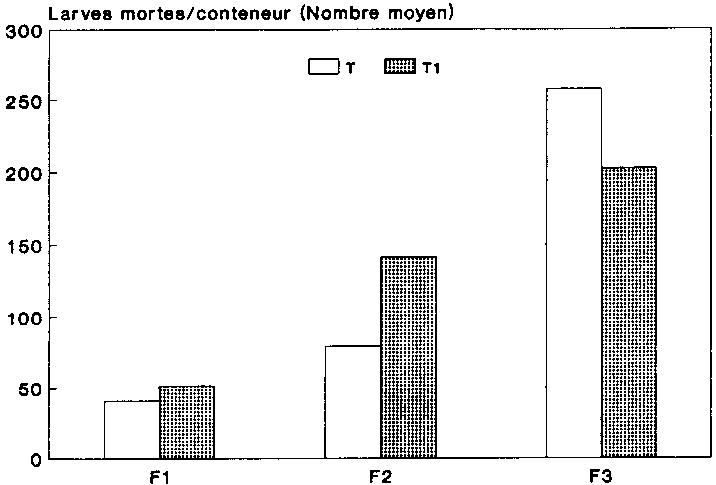
La figure 13 indique le nombre moyen des larves mortes au sein des g�n�rations filiales des deux variantes. La comparaison r�v�le que les chiffres diff�raient fortement au sein de la g�n�ration on F2. On a en effet d�nombr� ici dans les populations trait�es aux spores 550 larves mortes, alors que ce chiffre n'�tait que de 208 pour les populations-t�moins.
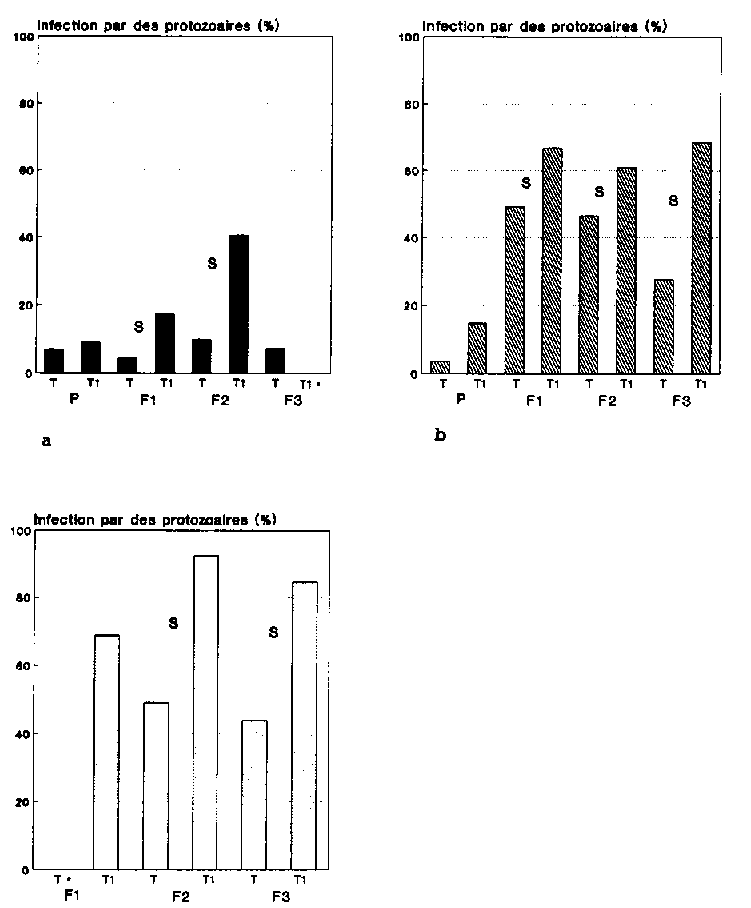
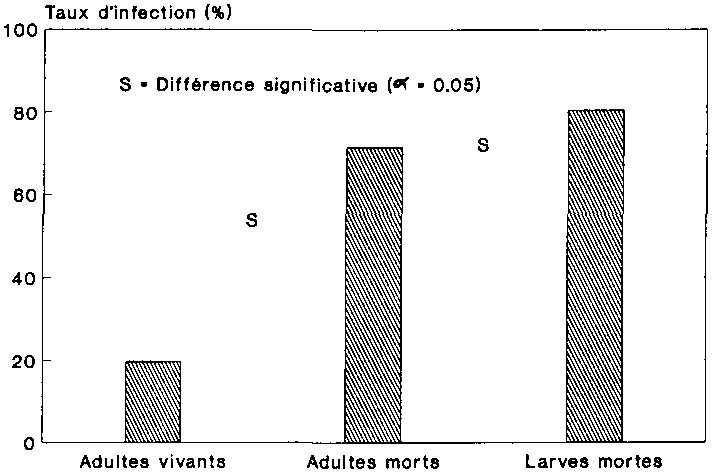
Les taux d'infection moyens des stades examin�s de l'anobie sont indiqu�s pour chacune des variantes sur la figure 14, a � c. La comparaison entre les taux d'infection des adultes vivants et ceux des adultes morts r�v�le que les col�opt�res morts �taient plus fr�quemment infest�s par des protozoaires que leurs cong�n�res vivants (cf. '-9 14, a et b). Ce r�sultat permet d�j� de conclure aux effets mortels des agents pathog�nes. S'agissant des stades de d�veloppement morts du ravageur, le nombre des larves infest�es par l'agent pathog�ne �tait beaucoup plus �lev� que celui des adultes (cf. fig 14, b et c). La comparaison statistique entre les taux d'infection observ�s chez les adultes et les larves infestes naturellement et les adultes, morts et vivants, ainsi que les larves mortes ayant subi un traitement, a r�v�l� qu'� l'exception de la g�n�ration P. les taux d'infection des divers stades trait�s �taient tous tr�s nettement sup�rieurs � ceux r�sultant de l'infection naturelle par les protozoaires (cf. fig. 14, a � c). Les diff�rences significative entre les stades de P. truncatus infestes naturellement et les stades trait�s ont �t� mises en relief sur la figure 14, a � c.
Le taux d'infection des adultes morts qui avaient subi un traitement auparavant �tait sup�rieur � 60 % pour les trois g�n�rations filiales (fig. 14 b). Parall�lement � ce degr� d'infection �lev�, les populations trait�es pr�sentaient des taux de mortalit� largement sup�rieurs � ceux du t�moin (cf. fig. 12). Ces r�sultats permettent de penser que chez les populations trait�es aux spores, l'infection de protozoaires a eu des r�percussions sur la mortalit� des col�opt�res. Aucun effet de ce genre n'a �t� observe au sein des populations infest�es naturellement. Le taux d'infection des adultes morts a baisse a une g�n�ration � l'autre, son r�le devenant ainsi de plus en plus n�gligeable dans la mortalit� constat�e (cf. fig. 14 b).
C'est dans la g�n�ration F2 des populations trait�es que l'infection des larves mortes par les protozoaires atteignait son plus haut niveau avec 92,5% (fig. 14 c). Le nombre remarquablement �lev� de larves mortes au sein de cette m�me g�n�ration est �galement � appr�cier dans ce contexte (cf. fig. 13). La plupart des larves sont mortes par cons�quent des suites de la maladie. Au-del�, on peut en d�duire que la forte r�gression observ�e au sein de F2 dans la dynamique des populations trait�es (cf. fig. Il) est � mettre pour l'essentiel au compte du taux de mortalit� �lev� affectant les larves. Les effets n'ont pas Et! les m�mes chez les populations exemptes de traitement aux spores, ce qui tient d'une part au fait que le nombre de larves mortes �tait beaucoup plus r�duit dans cette variante (cf. fig. 13), et que d'autre part les taux d'infection de ces animaux demeuraient en g�n�ral en de�� de 50 % (cf. fig. 14 c).
Fig.
14 : Comparaison des taux d'infection chez les Prostephanus
truncatus infest�s naturellement (T) et ceux trait�s aux spores
(T1) dans la premi�re g�n�ration et les g�n�rations filiales
1 � 3 (F1 � 3)
(valeur caract�ristique pour a = 0
05;* = pas d'observation d'individus)
a Infection des adultes vivants
b Infection des adultes morts
c Infection des larves mortes
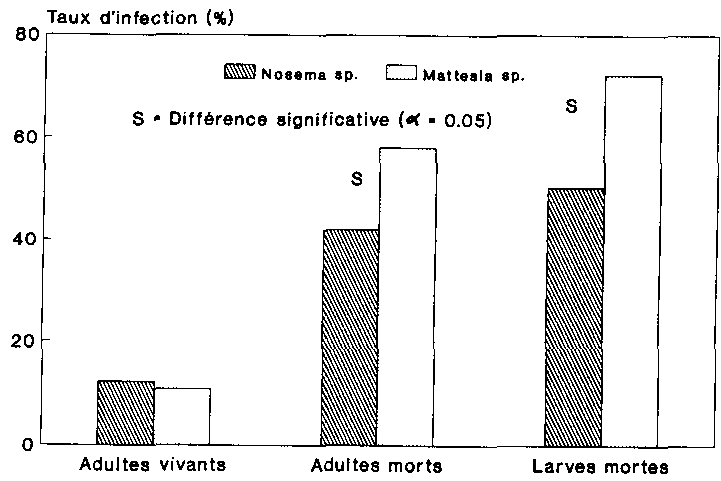
Pour terminer, nous examinerons s�par�ment les taux d'infection des deux protozoaires. Les figures 15 et 16 repr�sentent en les taux d'infection respectifs par Nosema sp. et Mattesia sp. des populations de P. truncatus pendant toute la dur�e de l'essai. Le test Chi2 utilise' qui vise � comparer les taux d'infection des animaux trait�s � ceux de leurs cong�n�res infestes naturellement, tenait compte ici des individus atteints d'une infestation mixte apparus dans les deux variantes.
L'esp�ce Nosema (tait beaucoup plus fr�quemment repr�sent�e au sein des populations trait�es que chez les populations non trait�es, et cela � tous les stades (fig. 15). Avec un taux d'infection maximal de 82,5 % chez les larves mortes, la microsporidie s'est de toute �vidence fort bien propag�e dans la population de l'h�te. Il en va tout autrement en ce qui concerne l'esp�ce Mattesia Les taux d'infection naturelle �taient ici dans tous les cas sup�rieurs aux taux d'infection obtenus � la suite de l'application � P. truncatus ri faut toutefois noter que cette �tablie par le test statistique, �tait uniquement significative au niveau des adultes morts (fig. 16). Le nombre total de sujets infect�s par l'esp�ce Mattesia atteignait au maximum 40 %, cette infestation pouvant d'ailleurs s'expliquer pour les deux variantes par l'infection naturelle de la g�n�ration parentale.
Le tableau 10 indique l'infestation naturelle moyenne par les protozoaires pr�sent�e par les P. truncatus utilises. Comme le montre ce tableau, les animaux non traites �taient assez fortement infestes par la n�ogr�garine, alors que l'esp�ce Nosema �tait pratiquement absente. L'esp�ce Mattesia disposait donc de conditions nettement plus favorables que Nosema sp. pour se propager dans la population.
La premi�re g�n�ration vivante de l'autre variante pr�sentait au contraire une infection plut�t uniforme, provoqu�e ici par les deux protozoaires. (Les animaux morts et infestes n'ont joue aucun r�le dans la propagation des protozoaires dans les populations du fait qu'ils ont �t� retir�s relativement t�t). L'addition de la pr�paration mixte aux spores, qui contenait beaucoup plus de spores de Nosema que de spores de Mattesia (cf. chap. 2.4.3), s'est traduite de toute �vidence par une propagation optimale de la microsporidie, alors que la n�ogr�garine n'a cause chez les poss�de toutefois aucun caract�re d'universalit� dans la mesure o� l'infection naturelle par les protozoaires repr�sentait dans cet essai une variable al�atoire.
Fig.
15 : Infection par Nosema chez les populations de Prostephanus
truncatus infest�es naturellement (t�moin) et chez celles
trait�es aux spores (traitement)
(Taux moyens aux divers stades de d�veloppement et pendant toute
la dur�e de l'essai)
Nous constaterons en r�sum� que l'�l�vation du taux de mortalit� observ�e chez les populations de P. truncatus trait�es aux spores est principalement due � l'infection par Nosema sp..
Tabl. 10 : Infection moyenne par Nosema sp.. et Mattesia sp. de la premi�re g�n�ration chez les populations de Prostephanus truncatus infest�es naturellement et chez les populations trait�es
|
Taux d'infection (%) |
|||
trait� |
non trait� |
|||
| vivants | morts | vivants | morts | |
| Mattesia sp. | 6,3 | 3,7 | 4,2 | 11 |
| Nosema sp. | 0,6 | 0 | 4,9 | 3,7 |
3.3.5.2 Inoculation pratiqu�e sur des Prostephanus truncatus
sains
Pour l'essai en conteneur, lequel comportait trois r�p�titions et avait pour objet d'observer la dynamique d'une population � la suite d'un traitement � une pr�paration mixte aux spores, on a utilis� 10 m�les et femelles sains de P. truncatus par verre. Le nombre de col�opt�res morts et vivants, ainsi que le nombre de larves mortes, ont �t� recenses respectivement au bout de 4, 8, 12 et 16 semaines. On a par ailleurs �tabli le taux d'infection des sujets traites aux spores selon la m�thode d�sormais connue.
La figure 17 montre la dynamique des populations saines et celle des populations trait�es aux spores de protozoaires � partir du nombre moyen d'adultes vivants. Au sein de la g�n�ration Fi, la descendance a augmente dans les deux variantes de mani�re relativement uniforme. Alors que dans la seconde g�n�ration filiale, les populations saines ont continue de se reproduire, donnant naissance � pr�s de 800 individus, on a enregistre dans la g�n�ration F2 trait�e un recul du nombre de col�opt�res, qui n'atteignait plus en moyenne que 250 animaux. Pour ce qui est de la g�n�ration F3, enfin, on a constat� une r�gression au niveau de la dynamique dans les deux variantes. Avec 100 individus seulement, on ne d�nombrait cependant plus chez les populations trait�es qu'un tiers des animaux pr�sents dans les populations saines. La r�gression, a la fois pr�coce et accentu�e, observ�e chez les populations de ravageurs trait�es, constitue d�j� un indice de l'effet exerc� par les spores de protozoaires.
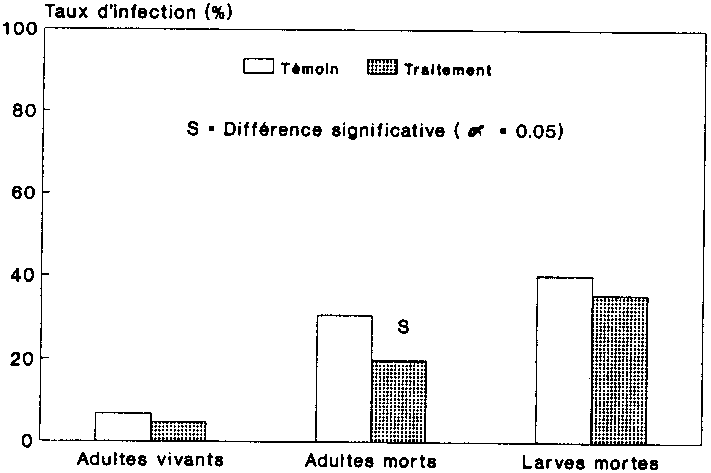
Le taux moyen de mortalit� des adultes de chaque variante indique qu'� l'exception de la premi�re g�n�ration, les col�opt�res trait�s des trois g�n�rations filiales sont morts en plus grand nombre que leurs cong�n�res non trait�s (fig. 18). Le test Chi� 2 n'a permis d'�tablir une diff�rence significative entre les variantes que pour la g�n�ration F2. La mortalit� enregistr�e dans les deux variantes est demeur�e faible, puisque le nombre global de col�opt�res morts atteignait tout au plus 25,5 %.
Fig.
18: Mortalit� des adultes de Prostephanus truncatus chez les
populations saines (t�moin) et chez les populations trait�es
aux spores (traitement)
(P = premi�re g�n�ration, F1 � F3 = g�n�rations filiales 1
� 3)
La figure 19 indique le nombre de larves mortes au sein des g�n�rations filiales. S'agissant de la g�n�ration F1, on a trouv� dans les deux variantes un nombre de larves mortes sensiblement �gal. Dans la seconde g�n�ration le nombre de larves mortes �tait deux fois sup�rieur � celui des populations saines, tandis que l'on observait la situation inverse pour la g�n�ration F3. On a en effet d�nombr� 258 larves dans les populations-t�moins, ce qui repr�sente 56 animaux de plus que les 202 larves mortes trouv�es chez les populations trait�es. Ces r�sultats sont indiff�rent.
En compl�ment de le dynamique et la mortalit� dans les deux variantes, on a �galement �tabli les taux d'infection des diff�rents stades des populations de P. truncatus trait�es aux spores. La figure 20 repr�sente les taux d'infection moyens pendant toute la dur�e de l'essai. Ce sont les larves mortes du ravageur qui, avec un taux de 80 %, �taient les plus fortement infest�es par l'agent pathog�ne. Les adultes morts �taient, eux, infestes � 70 % par des protozoaires, les P. truncatus vivants ayant subi l'infestation la plus minime, avec pr�s de 20 %. Le test Chi� a r�v�l� des diff�rences significatives entre les taux d'infection des divers stades de P. truncatus (cf. fig. 20).
Ces r�sultats d�montrent que l'application de spores de protozoaires a provoque dans les populations du ravageur de graves infestations. Bien que la maladie caus�e par les protozoaires se soit tr�s fr�quemment manifest�e chez les larves et les adultes morts, on n'a pas observe d'augmentation significative de la mortalit� des sujets trait�s par rapport aux sujets sains (cf. fig. 18). N�anmoins, vu que le nombre des individus qui se sont d�velopp�s sous l'effet des spores �tait nettement inf�rieur, nous allons examiner dans la suite plus en d�tail la multiplication dans les deux variantes.
Le tableau 11 met en opposition le nombre moyen d'individus compt�s dans les quatre g�n�rations, d'une part dans les populations saines, d'autre part dans les populations trait�es aux spores. Comme on peut le constater, il n'y a eu multiplication des P. truncatus trait�s dans aucune des trois g�n�rations filiales, puisque aussi bien le nombre d'individus d�cro�t au contraire d'une g�n�ration � l'autre. Sur les 544 sujets que comptait la g�n�ration F1, on n'en d�nombrait plus que 413 dans la seconde g�n�ration et 334 seulement dans la g�n�ration F3. Avec 895 sujets, les anobies saines se sont en revanche multipli�es jusqu'� la g�n�ration F2. Ce n'est qu'� la g�n�ration F3, qui comptait 729 individus, que l'on a constat� ici une r�gression dans la dynamique. Le recul pr�coce et tr�s marqu� de la dynamique, de m�me que les taux d'infection �lev�s dans les populations trait�es, permet d'en d�duire que la maladie caus�e par les protozoaires a entra�n� de toute �vidence une r�gression de la reproduction du ravageur.
Tabl. 11: Multiplication des populations de Prostephanus truncatus saines et de celles trait�es aux spores (Nombre moyen de larves et d'adultes morts respectivement au bout de 4, 8, 12 et 16 semaines)
| Populations | P* | F1* | F2* | F3* |
| saines | 20 | 613 | 895 | 729 |
| trait�es | 20 | 544 | 413 | 334 |
* P = premi�re g�n�ration F 1 � 3 = g�n�rations filiales 1
� 3)
L'examen des taux d'infection moyens r�v�le qu'aux stades morts de P. truncatus, l'esp�ce Mattesia �tait, et cela de mani�re significative, plus fr�quemment repr�sent�e que la microsporidie (fig. 21). La concentration de spores contenue dans la pr�paration de Mattesia sp. �tant inf�rieure � celle de Nosema sp., Mattesia sp. appara�t comme le plus virulent des deux protozoaires. Ce r�sultat est en contradiction avec le premier essai des populations. En effet, bien que la concentration de spores ait �t� la m�me dans les deux pr�parations, le nombre de sujets infest�s par Mattesia sp. �tait nettement inf�rieur � celui des sujets infest�s par la microsporidie. La r�ussite tr�s in�gale des essais d'infraction par Nosema sp. et Mattesia sp. pourrait s'expliquer par des rapports concurrentiels d'ordre g�n�ral.
On peut constater enfin que le traitement aux spores a permis, au cours des deux essais des populations, d'enregistrer un taux d'infection �lev� � la fois chez les adultes morts (plus de 60 %) et chez les larves mortes du ravageur (plus de 80 %). Le test Chi� a r�v�l� que les sujets morts �taient, de mani�re significative, plus fr�quemment infestes par les protozoaires que leurs cong�n�res vivants. On a donc pu mettre en �vidence les effets mortels de l'agent pathog�ne parmi toutes les populations de P. truncatus comportant des individus infestes par les protozoaires.
3.3.6 Contagiosit� des spores de protozoaires apr�s stockage
La long�vit� des spores de protozoaires constitue un facteur important du point de vue du stockage des insecticides biologiques. Ce type de pr�parations, qui contient des agents pathog�nes microbiens, doit �tre mis en formule et stabilise pour que ses propri�t�s insecticides ne soient pas alt�r�es dans des conditions de stockage normales (COUCH & IGNOFFO, 1981).
Afin de d�terminer la capacit� au stockage des spores de Nosema et de Mattesia on a conserve celles-ci durant des p�riodes plus ou moins longues et dans des conditions vari�es, avant d'en v�rifier la contagiosit� sur des larves de P. truncatus.
Stockage des spores dans des cadavres d'insectes
La conservation des spores dans des cadavres de l'h�te constitue une m�thode relativement simple et pourrait donc, le cas �ch�ant, �tre pratiqu�e par les paysans eux-m�mes Nous allons par cons�quent nous pencher dans la suite sur cette forme de conservation des spores, qui offre un int�r�t pratique �vident.
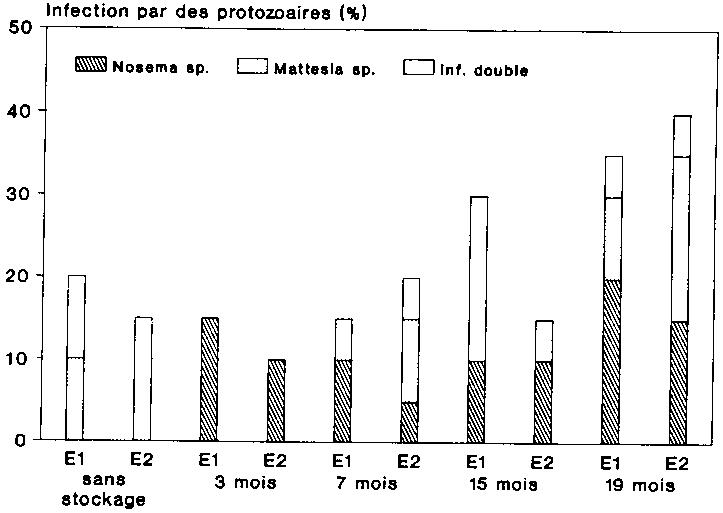
On a ainsi analyse le pouvoir infectieux des spores de protozoaires � la suite d'un stockage de 3, 7, 15 et 19 mois dans des P. truncatus morts, � une temp�rature ambiante de 28 "C. A l'issue des diverses par odes de stockage, on a broy� une partie des cadavres afin d'obtenir une poudre aux spores, � laquelle on �t� traites � chaque fois 20 larves de P. truncatus. Deux essais d'inoculation ont eu lieu, les verres de l'essai 2 contenant une quantit� de spores trois fois sup�rieure � ceux de l'essai 1 (cf. tabl. 1).
La figure 22 indique les taux d'infestation enregistr�s parmi les larves, r�partis entre Mattesia Nosema et les infections doubles. On notera en premier lieu que les larves de l'anobie ont �t� infect�es quelle que soit la variante consid�r�e. L'�ge des spores ne jouait manifestement aucun r�le du point de vue des r�sultats. Les spores de la pr�paration qui n'avait pas �t� stock�e ont provoque de 15 � 20 % d'infections au cours des deux essais, alors que l'infestation subie par les larves a �t� avec 35 % (essai 1) et 40 % (essai 2), la plus forte sous l'effet des spores conserv�es durant 19 mois.
L'inoculation a une triple quantit� de spores (essai 2) ne s'est traduite que pour deux variantes de stockage par un taux d'infection plus �lev� chez les larves, alors qu'au contraire les spores des trois autres variantes de l'essai 2 ont infect! moins de sujets qu'au cours de l'essai 1. L'augmentation de la concentration des spores n'a donc eu des incidences positives sur la contagiosit� chez les larves de P. truncatus que pour un nombre restreint de variantes.
A l'inverse des r�sultats obtenus dans les autres variantes, les taux d'infection �tablis chez les larves auxquelles on a inocule des spores stock�es durant 19 mois peuvent �tre consid�r�s comme relativement �lev�s (cf. fig. 22). Dans la mesure o�, aussi bien dans l'essai I que dans l'essai 2, la concentration de spores des pr�parations de cette variante �tait � proportion sup�rieure (cf. tabl. 1), les taux d'infection (�lev�s qui ont �t� constates sont beaucoup plus vraisemblablement dus � la quantit� des spores contenues dans la poudre qu'� l'�ge de ces m�mes spores.
S'agissent des r�sultats d'ensemble, on peut affirmer en outre que l'infection provoqu�e chez les larves par les spores stock�es plus de � mois {tait uniquement due a Mattesia et cela dans les deux essais. D'autre part, l'inoculation, dans le cadre de l'essai 2, d'une poudre aux spores fra�chement pr�par�e, a �t� exclusivement suivie d'une infection par la n�ogr�garine. alors que dans la m�me variante de l'essai 2 les deux protozoaires ont infeste un nombre �gal de larves. Pour toutes les autres variantes de stockage, l'inoculation de spores a entra�n� aussi bien une infection par Nosema sp. et par Mattesia sp. (cf. fig. 22). Ces r�sultats permettent de conclure que les deux types de spores �taient conservables pendant au moins 19 mois dans les cadavres des animaux sans perdre de leur pouvoir infectieux.
Conservation au frais et � l'�tat congel� de la pr�paration aux spores
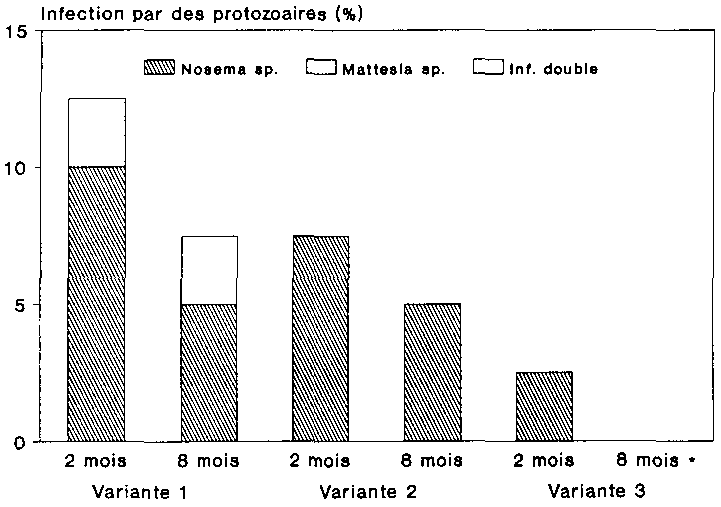
La dur�e de conservation des spores de Nosema sp. et de Mattes la sp. a {te �galement mise � l'�tude dans une pr�paration aux spores d�j� pr�te. Les pr�parations finies ont �t� stock�es � des temp�ratures de r�frig�ration (entre 4 et 10 �C) et de cong�lation (entre -12 et -22 �C). Une conservation � temp�rature normale (env. 28 �C dans le laboratoire) a servi de t�moin. La dur�e de stockage des pr�parations �tait respectivement de 2 et 8 mois. A l'issue du stockage, les pr�parations ont �t� inocul�es � 40 larves de P. truncatus par variante. Les taux d'infection obtenus chez des larves trait�es au moyen d'une quantit� de spores identique sont indiques sur la figure 23.
L'inoculation � des larves de P. truncatus de la pr�paration aux spores soumise � un stockage � temp�rature normale (variante 1) s'est traduite par une infection provoqu�e par les deux esp�ces de protozoaires, bien qu'apr�s 8 mois de conservation, Mattesia ne se soit plus manifest�e que sous forme d'infection mixte. Dans le cas du stockage au froid (variante 2) et de la cong�lation (variante 3), en revanche, l'infection d�cel�e chez les larves �tait uniquement due � Nosema L'absence d'infection par Mattesia sugg�re que les spores de cette esp�ce ont �t� tu�es aussi bien par la conservation eu froid que par la cong�lation.
La comparaison de l'infection par Nosema dans les trois variantes permet de constater que l'infestation moyenne la plus forte, qui avoisinait les 8 %, a �t� observ�e apr�s stockage de la pr�paration aux spores a une temp�rature de 28 'C (variante 1). Les spores de Nosema conserv�es au frais, quant � elles, ont infect� environ 6 % des larves, et apr�s conservation par cong�lation (variante 3), 1 % seulement des larves de P. truncatus pr�sentait une infestation par la microsporidie (cf. fig. 23). Le taux d'infection par Nosema sp. diminuait donc parall�lement � la temp�rature de stockage des spores. Cet effet a �t� constat� aussi bien � l'issue de la p�riode de conservation de 2 mois qu'apr�s celle de 8 mois, la p�riode de stockage prolonge se traduisant en g�n�ral par des taux d'infection moindres. Enfin, l'inoculation de spores conserv�es pendant 8 mois en �tat de cong�lation n'a provoque aucune infestation chez les larves de l'anobie.
Les r�sultats d'essais pr�sentes ici semblent indiquer qu'aussi bien la r�frig�ration ion et la cong�lation de la pr�paration aux spores entra�nent pour Mattesia sp. la perte de sa pathog�nocit�, puisque aucune infestation par la n�ogr�garine n'a �t� constat�e chez les larves de l'anobie. Les spores de Nosema en revanche, se sont av�r�es jusqu'� un certain point stockables � de telles temp�ratures. Ainsi qu'en t�moignent les taux d'infection moindres, la conservation des spores � une temp�rature ambiante voisine de 28 �C s'est �galement av�r�e pour ce protozoaire plus fiable que le stockage au froid ou par cong�lation.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}