Biocontrol with Teretriosoma nigrescens
Recherches sur l'influence de Teretriosoma
nigrescens Lewis (Col.: Histeridae) sur le
développement des populations de Prostephanus
truncatus (Horn) (Col.: Bostrichidae) dans
des conditions semi-pratiques au Togo.
Ecology and biological control of the Larger
Grain Borer in Mexico and Honduras
Discussion
- third session
Helbig, J. *; Laborius, G.-A. ** et Schulz, F.A.
* * TU, Berlin and ** GTZ, Hamburg
RESUME
L'influence du prédateur Teretriosoma nigrescens Lewis (Col.: Histeridae) sur le développement des populations de Prostephanus truncatus (Horn) (Col.: Bostrichidae) sous des conditions climatiques togolaises a été étudiée avec un essai sous des conditions semi-pratiques. Les expériences ont été réalisées dans une cage (dimensions: 9 m x 5 m x 2,5 m), garnie d'un grillage en acier spécial, sur le terrain du Service de la Protection des Végétaux, à Lomé-Cacaveli, au Togo. Les 6 greniers à maïs ont été confectionnés sur le modèle traditionnel, c'est-à-dire que les épis y ont été stockés en spathes. Avant d'être isolés dans les 6 cellules de la cage, les épis de maïs ont été exposés durant plus de 4 mois aux infestations naturelles des ravageurs des denrées stockées. Dans trois des 6 greniers on a placé 2000 adultes du prédateur, tandis que trois autres greniers servaient au contrôle. Au cours des 8 mois qu'a duré l'expérience, T. nigrescens a pu réduire la population de P. truncatus de 80,6 % en moyenne, les pertes en matière sèche et les dommages causés au grain d'environ 40 % par rapport au contrôle. On n'a pu constater aucun problème d'adaptation du T. nigrescens aux conditions climatiques togolaises.
INTRODUCTION
A la suite de l'introduction de Prostephanus truncatus (Horn) (Col.: Bostrichidae) en Afrique, une comparaison générale a permis de constater que les partes provoquées par ce ravageur sur le maïs stocké en Afrique étaient beaucoup plus élevées que celles qu'il causait dans son habitat d'origine, l'Amérique centrale. Il s'agissait dans la première phase du projet GTZ de prouver le bien-fondé de ces observations à l'aide de données et d'identifier les causes de cette différence. C'est ainsi que l'on a enquêté au Costa Rica pour savoir si le ravageur y avait des antagonistes, les recherches s'orientant en l'occurrence aussi bien vers les micro-organismes que vers les insectes (Schulz et al.1987). On a trouvé Teretriosoma nigrescens Lewis (Col.: Histeridae), un prédateur sur lequel on fonde de grands espoirs et qui présente, comme l'ont révélé les essais, une haute spécificité. C'est ainsi qu'il s'oriente à la recherche de proies sur la phéromone d'aggrégation émises par P. truncatus (Boeye 1988; Boeye et al. 1992). Les expériences effectuées ont permis d'établir l'efficacité du prédateur (Boeye 1988; Leliveldt et al. 1989; Rees 1987, 1985). Avant la libération de l'organisme utile au Togo, les responsables togolais ont demandé à ce que l'on vérifie encore une fois son rendement dans les conditions spécifiques au pays.
MATERIELS ET METHODES
Les expériences ont été effectuées dans des cages munies d'un grillage d'acier spécial et placées sur le terrain du Service de la Protection des Végétaux, à Lomé-Cacaveli.
Greniers expérimentaux:
Les 6 greniers expérimentaux, du type «Ebli-va», traditionnel,
ont été érigés en août 1989 sur une plate-forme de béton (6
m x 10m) préparée à cet effet. Le châssis. qui reposait sur
quatre pieds (80 cm de haut), a été construit en bois de
l'espèce acajou, la plate-forme elle-même (120 cm de long sur
120 cm de large) étant constituée d'essences communément
utilisées par les paysans. Les branches étaient fixées par un
bandeau tressé. L'engrangement du maïs (env. 200 kg d'une
espèce locale à grains jaunes par grenier) a eu lieu les 23 et
24 août. Le maïs a ensuite été recouvert du toit de paille
typique. Les 15 et 16 novembre, les épis de maïs ont été
sortis des greniers, systématiquement mélangés, puis les
greniers ont été reconstitués. Cette mesure avait pour objet
d'assurer dans tous les greniers une répartition homogène des
insectes nuisibles.
Cage expérimentale:
La cage grillagée (5 m de large, 9 m de long, 2,50 m de haut)
était divisée en deux moitiés dans le sens longitudinal,
chaque moitié comprenant 4 cellules, voisines les unes des
autres. Il s'agissait de 3 cellules expérimentales (2,50 x 2,50
m) et d'une entrée, faisant office de sas ( 1,50 m x 2,50 m ).
Les deux moitiés ne communiquaient pas entre elles. A
l'intérieur de chacune des moitiés, les différentes cellules
étaient reliées entre elles par une porte. La même moitié de
cage a servi aux trois répétitions du traitement. Le cadre de
la cage a été fabriqué en acier plat et enduit d'un produit
anti-corrosion. Le grillage en acier spécial (largeur de
mailles: 0,5 mm) a été tendu entre les montants en acier plat
et, pour en assurer l'étanchéité absolue, traité avec une
matière spéciale à étancher. L'écoulement de l'eau se
faisait séparément pour chacune des deux moitiés de la cage au
moyen de siphons aboutissant respectivement dans deux
collecteurs.
Introduction de T. nigrescens:
Le transfert des sujets adultes de T. nigrescens de
l'Institut biologique fédéral (BBA) de Berlin à Lomé a eu
lieu le 17 novembre 1989. C'est le 18 décembre 1989 que les
adultes du prédateur, c'est-à-dire 2000 par répétition, ont
été introduits dans les cages. Le prédateur est originaire du
Costa Rica, où les coléoptères ont été collectés en
novembre 1988 dans des champs de maïs à l'aide de pièges à
phéromones (Trunc-call 1). 5 à 6 générations d'entre eux ont
été observés au BBA dans des conditions de Quarantaine. On les
a fait se reproduire et on les a examinés afin de déceler sur
eux d'éventuels pathogènes. Les coléoptères ne présentaient
aucun pathogène ni antagoniste.
Échantillonnage:
Les premiers échantillonnages de maïs ont été effectués en
août 1989, aussitôt après la mise en stock. Le prélèvement
du maïs dans les greniers a été réalisé selon la méthode
décrite par Pantenius (1987). La surface du grenier a été
fictivement divisée en 4 quarts. On a prélevé de chacun de ces
quarts 25 épis, dont 15 provenaient de la périphérie et 10 de
la zone intérieure du grenier. Après avoir effectué deux
autres prélèvements de 100 épis, avec un intervalle d'un mois
entre les prélèvements, on a interrompu le processus. Pendant
cette interruption, les épis ont été de nouveau mélangés,
les greniers reconstitués et la cage placée au-dessus des
greniers. Après la fermeture de la cage, maïs avant
l'introduction de T. nigrescens, on a recommencé à
prélever régulièrement des échantillons. Afin d'empêcher
l'organisme utile de s'échapper, on a, avant de procéder à
leur évaluation, fumigé ces échantillons durant au moins une
semaine au PH3, dans une caisse à châssis placée dans
l'entrée de la cage, ce qui a également eu pour effet
d'anéantir la totalité des insectes.
Evaluation:
Pour l'évaluation, on a procédé à l'analyse individuelle des
épis. On a établi pour chacun d'entre eux le nombre des grains
infestés et le nombre et l'espèce des insectes nuisibles qui
s'y trou-vaient. Le tamisage des échantillons a été effectué
au moyen de tamis à mailles de 3,15 et de 0,4 mm. La fraction de
grain qui est demeurée dans le tamis de 3,15 mm et la farine de
forage, la fraction qui est passée au travers des mailles de 0,4
mm, ont été pesées. Après l'évaluation des 100 épis, on a
pesé l'ensemble des trois fractions (grains intacts, grains
infestés et farine de forage) et compté les grains intacts à
l'aide d'un compteur de grains (Numigral). Pour effectuer le
calcul des dommages et des pertes, on a appliqué la méthode du
comptage et du pesage.
% de dommages = (B / A) x 100
% de pertes = [(ExB)- (CxD)]/(ExA) x 100
A = nombre de grains totals
B = nombre de grains intacts
C = nombre de grains infestés (y compris les grains entièrement
détruits)
D = poids des grains intacts
E = poids des grains infestés et des débris de grains
RESULTATS
1. Développement des populations de P. truncatus dans le grenier à maïs
Le développement des populations de P. truncatus dans les deux variantes utilisées pour les expériences a été très inégal. Alors que l'efficacité du prédateur ne s'est fait sentir que de manière diffuse au cours des 3 premiers mois, seuls les greniers non traités ont connu un accroissement sensible de la population de P. truncatus, tandis que les populations du grenier traité présentaient une nette stagnation (Fig. 1).
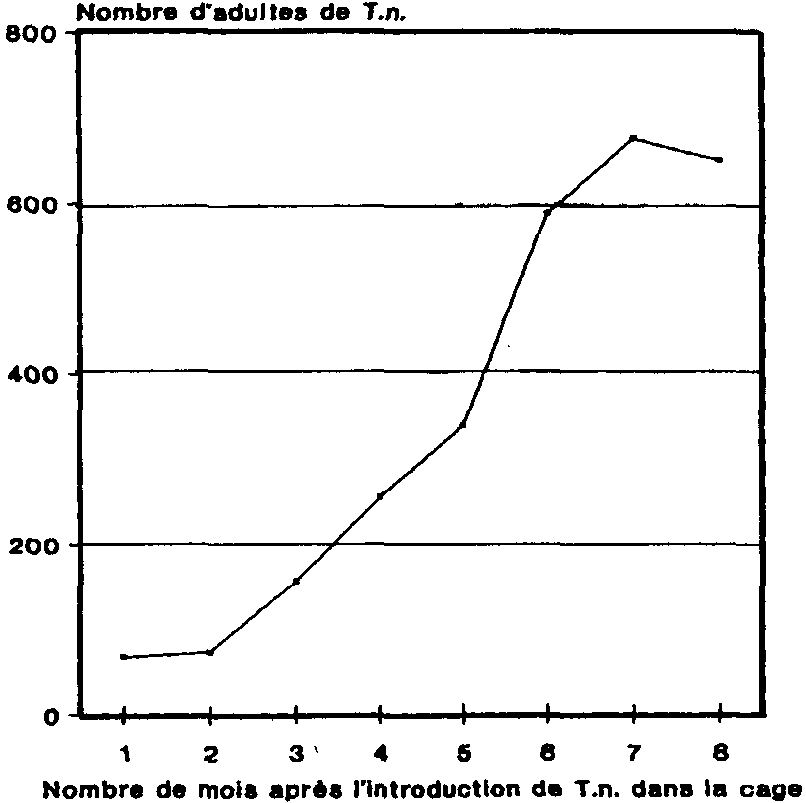
2. Développement de la population de T. nigrescens dans le grenier à maïs
Du 2ème au 7ème mois, le nombre des T. nigrescens a été en augmentation constante, pour atteindre un maximum d'env. 680 coléoptères pour 100 épis au bout du 7ème mois. Au cours du 8ème mois, on a observé un léger recul du nombre des T. nigrescens, ce qui semble indiquer que le prédateur avait acquis à ce niveau sa densité maximale et que, ayant quitté les épis de maïs, il se tenait désormais dans la zone libre de la cage.
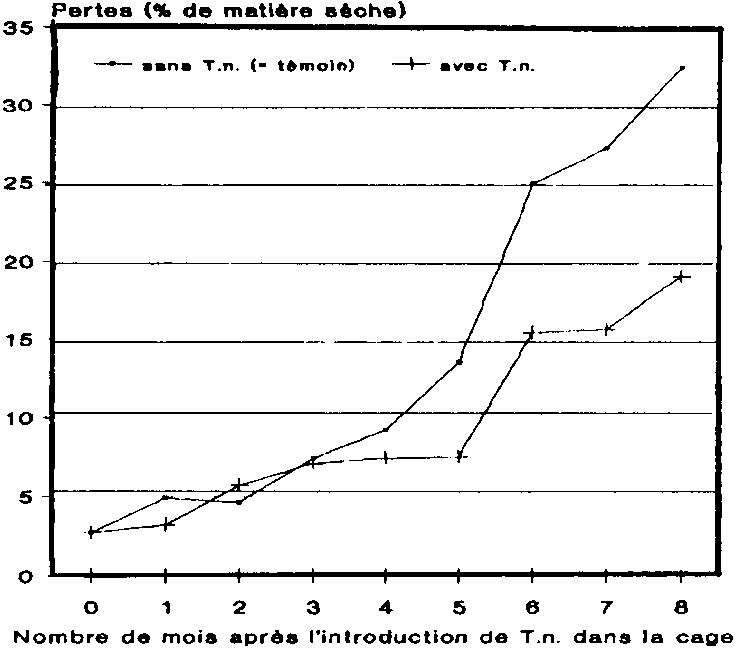
3. Evolution des pertes dans les greniers à maïs
Au cours des premiers mois gui ont suivi l'introduction du prédateur, l'évolution des courbes de pertes a été pratiquement identique dans les deux variantes. A partir du 4ème mois d'essai, les pertes enregistrées dans les greniers-témoins ont augmenté de manière beaucoup plus sensible que dans les greniers où l'on avait placé T. nigrescens. Au bout de 8 mois de stockage, les pertes enregistrées dans les greniers sans prédateur, qui se situaient à env. 32 % en valeur absolue, dépassaient ainsi de 40 % environ celles enregistrées dans les greniers avec prédateur qui, pour leur part, avoisinaient 19 % en valeur absolue. Ces chiffres démontrent clairement, à la lois les effets réducteurs de T. nigrescens sur les populations de P. truncatus, maïs également la réduction de pertes qui en découle. Il conviendra d'étudier dans une publication ultérieure les fluctuations de population des autres espèces de ravageurs, de même que leur influence sur l'évolution des pertes.
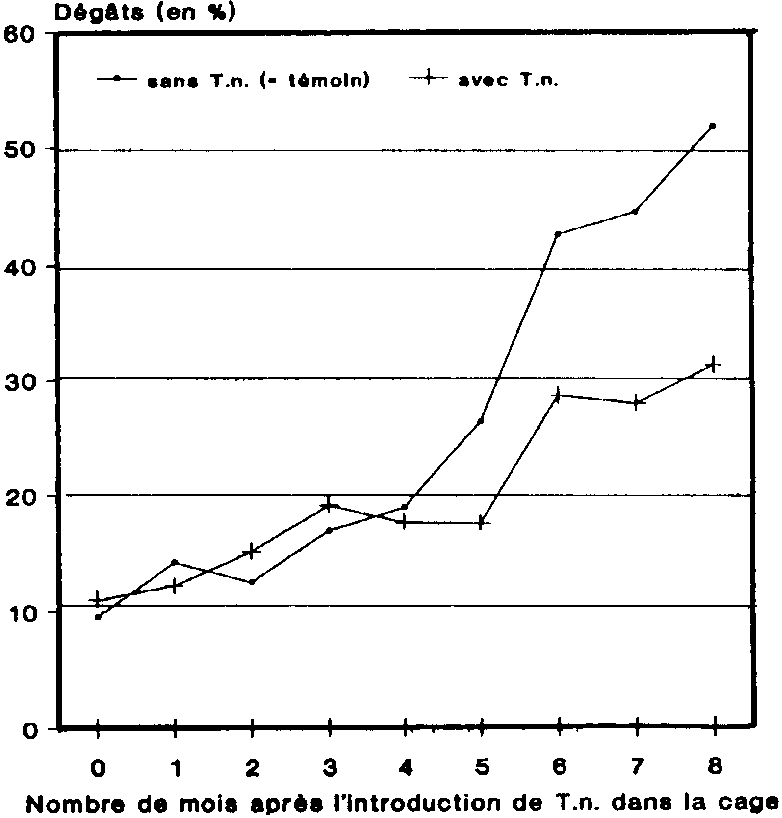
4. Evolution des dégâts au cours du stockage du maïs dans les greniers
Durant les premiers mois, on a constaté dans les deux variantes un léger accroissement des dommages. A partir du Sème mois, les dégâts enregistrés dans les greniers-témoins ont été en forte augmentation. Dans les greniers où se trouvait le prédateur, les dommages n'ont commencé à s'aggraver qu'un mois plus tard et n'ont pas augmenté aussi fortement. Au bout de 8 mois, les dommages causés dans les greniers sans T. nigrescens s'élevaient à env.52 % en valeur absolue. Dans les greniers où se trouvait le prédateur, les dommages, qui étaient de 40 % inférieurs, se limitaient à env. 31 % en valeur absolue.
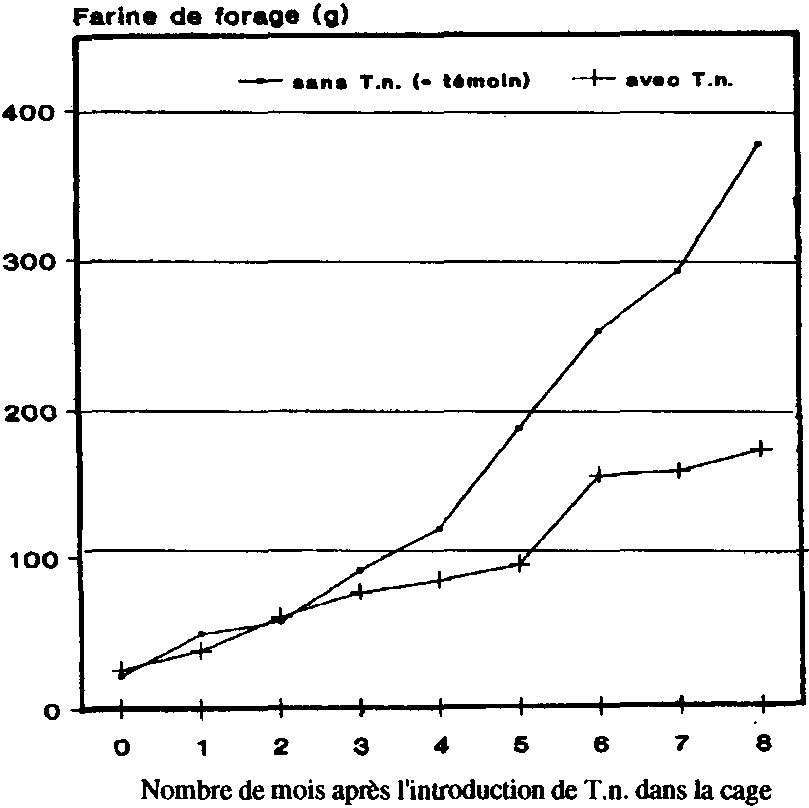
5. Production de farine de forage dans les greniers à maïs
La production de farine de forage a évolué de manière analogue à l'accroissement de la population de P. truncatus. Alors qu'au début de l'expérience on n'avait pas noté de différences vraiment tangibles entre les traitements, les différences sont apparues de plus en plus nettement au fur et à mesure que les essais se poursuivaient. Au bout de 8 mois, le poids de la farine de forage trouvée dans les greniers-témoins, à savoir 375 g env., était plus de deux fois supérieur (pour 100 épis) à celui enregistré dans les greniers où se trouvait le prédateur, où l'on a trouvé env. 170 g de farine de forage.
CONCLUSIONS
Au cours de ces expériences, le développement de la population de Prostephanus truncatus a pu être limité de façon efficace par le prédateur Teretriosoma nigrescens. Durant la période d'observation de 8 mois, cette population n'a que faiblement progressé. Comparé à celui des greniers-témoins, le nombre des P. truncatus adultes était de 80,6 % inférieur en présence du prédateur. Ce résultat a pu être obtenu en dépit du fait que l'introduction du prédateur dans les cages n'est intervenue qu'à une époque relativement tardive. Au cours d un essai effectué en conteneur, Rees (1985) a constaté que le développement ultérieur de la population du ravageur était étroitement fonction du décalage temporel entre le début de l'infestation par P. truncatus et l'apparition de T. nigrescens. En comparaison des résultats obtenus par Boeye (1988) au Costa Rica sur la base de prélèvements d'échantillons dans des greniers à maïs, le développement de la population de P. truncatus au cours de l'essai qui nous intéresse a été moindre. Dans les recherches menées par Boeye (1988), on a trouvé à l'issue d'une période de stockage allant de 5 à 7 mois entre 16 et 18 P. truncatus par épi , et cela malgré la présence de T. nigrescens. Il faut ce pendant noter que le nombre des prédateurs ne dépassait pas 2 ou 3 individus par épi, alors qu'il était de 6 à 7 au cours de la présente expérience, ce qui pourrait expliquer le développement plus accentué de la population du Grand Capucin du Maïs dans les recherches menées par Boeye (1988).
Lors de la mise en oeuvre de T. nigrescens on s'est efforcé de respecter le rapport numérique de 1 T.n. pour 10 P.t., qui s'était avéré optimal dans le cadre des recherches effectuées par Leliveldt et al. (1989). Sur la base des résultats d'essai obtenus l'année précédente au même emplacement, on a calculé le nombre de P. truncatus susceptibles de se trouver dans le grenier au moment de la mise en oeuvre du prédateur, et divisé ce nombre par le facteur 10, ce qui a donné approximativement le nombre de 2000 T. nigrescens par grenier. Il est toutefois permis de penser que cette valeur était trop élevée, et qu'il y a eu au cours des mois suivants un resserrement plus accentué du rapport numérique entre le prédateur et le ravageur qu'il ne l'a été dans les conditions pratiques choisies au Costa Rica par Boeye (1988). Dans des conditions pratiques, ce rapport se régularise par ailleurs lentement par immigration dans le grenier de l'ensemble des espèces et par multiplication à l'intérieur de celui-ci, ce qui n'a pas pu être observé en l'occurrence dans des conditions semi-pratiques, et cela pour des raisons techniques liées au type d'essai choisi. Si les résultats de ces essais suscitent un optimisme certain, ii faudra cependant se garder de les surestimer en voulant les appliquer sans la moindre critique.
Comme en témoignent les chiffres, le développement de la population du prédateur dans les cages n'a posé aucun problème. Le nombre des coléoptères adultes a pratiquement décuplé en l'espace de 6 mois. Il semble donc qu'il n'y ait pas eu de difficultés d'adaptation aux conditions climatiques. La stagnation enregistrée dans l'accroissement de la population de T. nigrescens à la fin de la période d'observation pourrait être due au rapport prédateur-proie, devenu alors trop étroit, ce que l'on avait pu observer un certain nombre de fois déjà au cours d'expériences en laboratoire (Boeye 1988; Leliveldt et al. 1989; Rees 1985). Lorsque le rapport prédateur-proie devient trop restreint et que la nourriture vient ainsi à manquer, ii y a de toute évidence apparition chez T. nigrescens d'un mécanisme de régulation, dans lequel ii faut voir un facteur important de conservation de l'espèce (cannibalisme).
Il se peut que d'autres possibilités d'interprétation s'offrent à nous lorsque nous serons en possession des chiffres concernant l'évolution des populations des autres espèces d'insectes présentes dans le grenier. Du fait que T. nigrescens se nourrit en partie de la descendance de ces espèces, ce comportement constitue dans la pratique une sorte de «tampon», dont l'intégration et l'interprétation quant au nombre de prédateurs présentent encore quelque difficulté. L'influence de T. nigrescens sur le développement de la population de P. truncatus se reflète également dans l'évolution de l'état des pertes. Bien que la population du ravageur ait pu se développer sans entraves 4 mois durant, les pertes, tout comme les dommages, enregistrées à la suite de l'introduction du prédateur, étaient inférieures de 40 % à celles constatées dans le grenier-témoin. Ceci est en l'occurrence très nettement dû à l'absence d'une population élevée de P. truncatus du point de vue numérique. Ces résultats permettent d'espérer que T. nigrescens permettra également dans la pratique de réduire considérablement les taux de population. On peut partir du principe qu'en milieu naturel, les effets de l'action exercée par le prédateur sur les populations de ravageurs se font sentir beaucoup plus tôt et que l'on peut obtenir un taux de réduction efficace, même à partir d'une population de T. nigrescens à l'origine peu nombreuse.
Ce que l'on n'a pas encore pu évaluer jusqu'à présent, c'est le rôle que joue T. nigrescens par rapport aux populations de P. truncatus dans la nature, loin des greniers à maïs. Si le prédateur était là encore en mesure de causer une réduction des populations de ravageurs, le potentiel d'infestation des deux denrées de stockage que sont le maïs et le manioc s'en trouverait d'emblée considérablement réduit.
BIBLIOGRAPHIE
Boeye, J.(1988) Autökologische Untersuchungen zum Verhalten des Großen Kornbohrers Prostephanus truncatus (Horn) (Coleoptera; Bostrichidae) in Costa Rica. Dissertation, Kiel, 195pp.
Boeye, J., Laborius, G. -A. and Schulz, F. A. (1992) The response of Teretriosoma nigrescens Lewis (Col.: Histeridae) to the pheromone of Prostephanus truncatus (Horn) (Col.: Bostrichidae). Anz. Schädlingskde., Pflanzenschutz, Umweltschutz, in press.
Leliveldt, B. and Laborius, G.-A. (1989) Effectiveness and specificity of the antagonist Teretriosoma nigrescens Lewis (Col.: Histeridae) on the Larger Grain Borer (Prostephanus truncatus (Horn) (Col.: Bostrichidae). pp 87-102 in Markham, R.H. and Herren, H.R. (Eds.), Biological control of the Larger Grain Borer, Proceedings of an IITA/FAO Coordination Meeting, Cotonou, Rep. of Benin. 2-3 June 1989
Pantenius, C.U.(1987) Verlustanalyse in kleinbäuerlichen Maislagerungssystemen der Tropen, dargestellt am Beispiel von Togo. Dissertation, Kiel, 249 pp.
Rees, D. P. (1985) Life history of Teretriosoma nigrescens Lewis (Coleoptera; Histeridae) and its ability to suppress populations of Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 21, 115- 118.
Rees, D. P. (1987) Laboratory studies on predation by Teretriosoma nigrescens Lewis (Coleoptera; Histeridae) on Prostephanus truncatus (Horn) (Coleoptera; Bostrichidae) infesting maize cobs in the presence of other maize pests. J. Stored Prod. Res. 23, 191 -195.
Schulz, F.A. and Laborius, G.-A. (1987) Strategy for Bio-integrated Control of Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae). pp 497-503 in Donahaye, E. and Navarro, S. (Eds.), 4th Int. Work. Conf Stored Prod. Prot., Tel Aviv, Israel, Sept. 21-26, 1986.
Ecology and biological control of the Larger Grain Borer in Mexico and Honduras
R.M. Rios Ibarra *, R.H. Markham **, P. Novillo Rameix *** and
V.F. Wright ***
* Colegio de Postgraduados, Mexico; ** IITA (BCP), Benin and ***
Escuela Agricola Panamericana, Honduras
INTRODUCTION
The severe losses suffered by stored maize in Africa, following the introduction of the larger grain borer, Prostephanus truncatus (Col.: Bostrichidae), and difficulties encountered by conventional, pesticide-based campaigns have led to widespread interest in the potential of biological control for management of this pest. However, the special characteristics of the larger grain borer, both as a «storage insect» which nevertheless occurs widely in natural habitats and as an «exotic pest» which sometimes causes severe damage in its area of origin, call for a cautious approach to classical biological control of this insect (Markham, 1990). Successful use of biological control as part of a broad strategy to control P. truncatus in Africa will depend on a good understanding of the factors which affect the incidence of the pest in its area of origin, in Mexico and Central America. A basis for such an understanding has been laid by the ecological observations of Boeye (1988) in Costa Rica, leading to the identification of a predator, Teretriosoma nigrescens (Col.: Histeridae), which shows some promise as a biological control agent (Leliveldt and Laborius, 1990; Rees 1987). However, a previous meeting, reviewing information so far available on P. truncatus and its natural enemies (Markham and Herren, 1990), concluded that additional information was needed on the importance of natural enemies in population regulation of P. truncatus and that factors leading to pest outbreaks in the Neotropics (despite the presumed presence of co-adapted natural enemies) needed to be investigated.
Studies initiated by the Biological Control Program (BCP) of the International institute of Tropical Agriculture (IITA) have sought to investigate these and related aspects of P. truncatus ecology at sites in Mexico (in collaboration with the Colegio de Postgraduados) and in Honduras (with the Escuela Agricola Panamericana, EAP). Some preliminary results of the first season's study, with emphasis on the
The main objective of this first phase of the studies was to observe the basic population dynamics of P. truncatus through a full storage season under "typical" rural storage conditions. The main study was supplemented, on the one hand, by wider surveys of the incidence of P. truncatus and related species and, on the other hand, by preliminary field and laboratory studies to elucidate factors influencing the population dynamics of P. truncatus at the main study site. Particular attention was given to investigation of possible sources of infestation outside the grain storage system and the importance of active migration as revealed by flight traps.
MATERIALS AND METHODS
Mexico
Based on reports in the literature and preliminary surveys of farmers' stores, study areas in both Mexico and Honduras were selected where maize is stored on-farm, using traditional methods, and where P. truncatus occurs consistently. The main study site in Mexico is at Chilcuautla, at an altitude of 1880 m, approximately 100 km directly north of Mexico City, in Hidalgo State. The study site consists of a small valley, with intensive arable cultivation (mostly irrigated maize on the valley floor and an adjacent village. Surrounding hillsides mainly support semi-natural vegetation of dry scrub, dominated by leguminous shrubs (especially Prosopis sp.), Cactaceae and other xeromorphic plants.
Three experimental cribs were constructed of local materials in the farm-yard of a small-scale maize farmer. Maize, grown (rainfed) on nearby uplands, was purchased from the same farmer and harvested by hand in late October 1989. The maize was stored in the cribs following normal local practice, that is "on the cob", without husks. The maize cribs had open lattice walls of local "bamboo" and were divided internally with the same material to form 32 compartments, accessible from the top. Each compartment was filled with approximately 100 maize cobs. At the time of loading each cob was inspected and the presence of any insects or insect damage recorded. Samples were subsequently collected from the cribs every two weeks until September 1990. On each occasion, one section (preselected at random) from each of the cribs was emptied and a sample of sixty cobs collected in a stratified random manner (20 each, from top, middle and bottom). The sampled cobs were placed carefully and rapidly into plastic bags as they were removed from the cribs and taken back to the laboratory for assessment. Cobs not used for sampling were replaced in the section of the crib from which they had been taken so as to maintain the integrity of the mass of grain.
In the laboratory, cobs were shelled, sieved and all adult insects collected from the grain. The grain was weighed and subsamples taken for determination of moisture content (by oven drying) and damage/ weight loss (count and weigh method, based on a sample of 1000 grains). Another subsample (1000 g) was incubated in the insectary (27°C, 70% relative humidity) and emergences of adults of primary pests and their parasitoids were assessed after one, two, three and four weeks. Data were corrected for sample size (adult counts) and grain moisture content (adults and emergences); population densities are thus expressed here per 1000 g of dry grain.
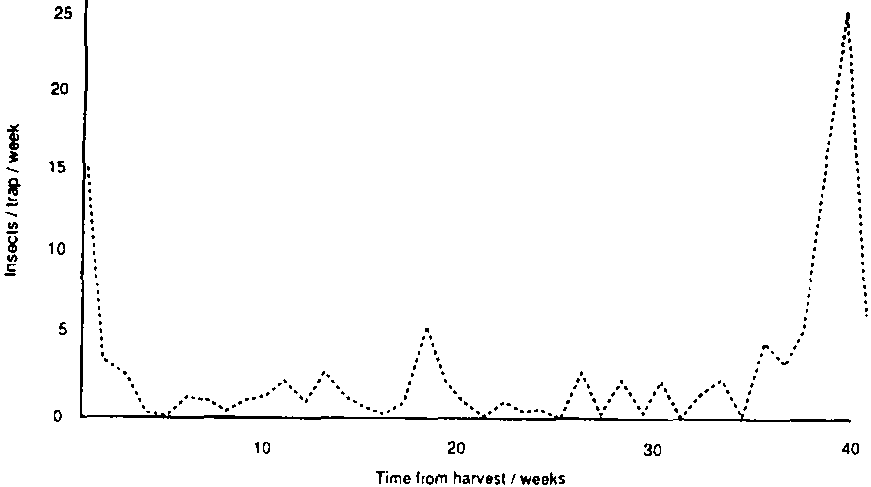
Flight activity in the immediate vicinity of the cribs was assessed using three unbaited, flat sticky traps (c.30cm x 30cm). These were hung inside, below the roof, of each of the three experimental stores. Traps were inspected weekly to count the number of adults of each species. In addition, four delta-type sticky traps (Trécé Pherocon II), baited with P. truncatus T1/T2 aggregation pheromone (from Natural Resources Institute, UK), were placed at increasing distances from the experimental stores. The nearest (at c. 200m) was in the village, the next on the edge of the village's maize cultivation and the furthest two (at 1.5 and 2km away) in «natural» vegetation further up the valley. Traps and lures were changed weekly and the number of P. truncatus and T. nigrescens adults recorded.
Honduras
The study in Honduras is based on the valley of Zamorano, at an altitude of 800m, some 35km south-east of Tegucigalpa. The floor of the valley is under intensive cultivation, mostly rainfed maize, though including other cereals, horticultural and tree crops. The experimental store is sited on the EAP's large-scale farm in the centre of the valley; however, additional samples are being collected in more traditional situations in nearby villages in the same valley, as well as in the valley of Moroceli, altitude 600m, approximately 20km away. The remaining natural vegetation in both valleys consists of dry woodland, dominated by leguminous trees and shrubs, with moister woodland along permanent water courses, especially in the Zamorano valley. The watershed between the two valleys, reaching 1400m, is dominated by Quercus spp at lower altitudes and Pinus oocarpa higher up, though with extensive areas of cultivation and pasture.
The experimental store at this locality, following local practise, was a wooden shed in which the maize was stored, with husks intact, in stacks on the floor. Samples (each of 180 cobs) were taken from nearby maize fields in December, January and early February, 1990. The maize was harvested in late February 1990 and stacked so as to form three blocks within the store. Each block, approximately 4.5m long x 0.5m wide, was divided into 14 compartments with wire mesh screens, each compartment containing 100 cobs. One compartment from each block was sampled every three weeks after storage and sampling will continue until December 1990. Samples consisted of 10 cobs each, from top, middle and bottom of each compartment. Laboratory procedures and data gathered were the same as in Mexico.
At this site, flight traps were placed at 2km intervals along a transect, following the main road from Zamorano to Moreceli. Although trap catches of P. truncatus and T. nigrescens were checked weekly, the pheromone lures were only changed every tour weeks, so data for these periods are grouped together,
RESULTS
In Chilcuautla, Mexico, it was estimated (mainly on the basis of damage symptoms) that at the time of harvest 0.6% of the cobs were infested by P. truncatus 0.5% by Sitotroga cerealella (Lep.: Gelechiidae) and 0.2% by Sitophilus ?zeamais (Col.: Curculionidae). The grain moisture content at harvest was approximately 16%. At Zamorano, Honduras, in contrast, no infestation of maize in the field by P. truncatus was detected, despite the presence of this species in the pheromone-baited flight traps and despite the relatively late harvest, at 13.5% moisture content. Field infestation by Sitophilus zeamais averaged 0.5 insects per cob in December (when moisture content was 16%) and had approximately doubled by the time of harvest.
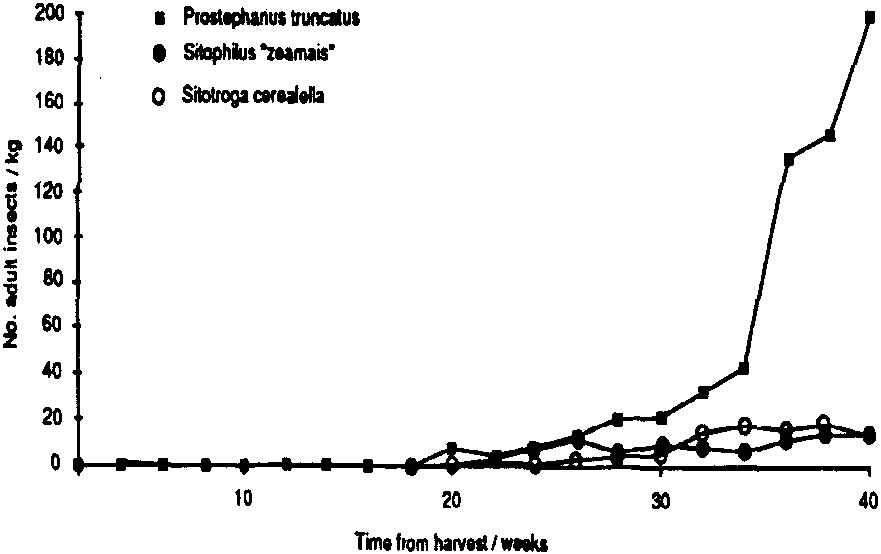
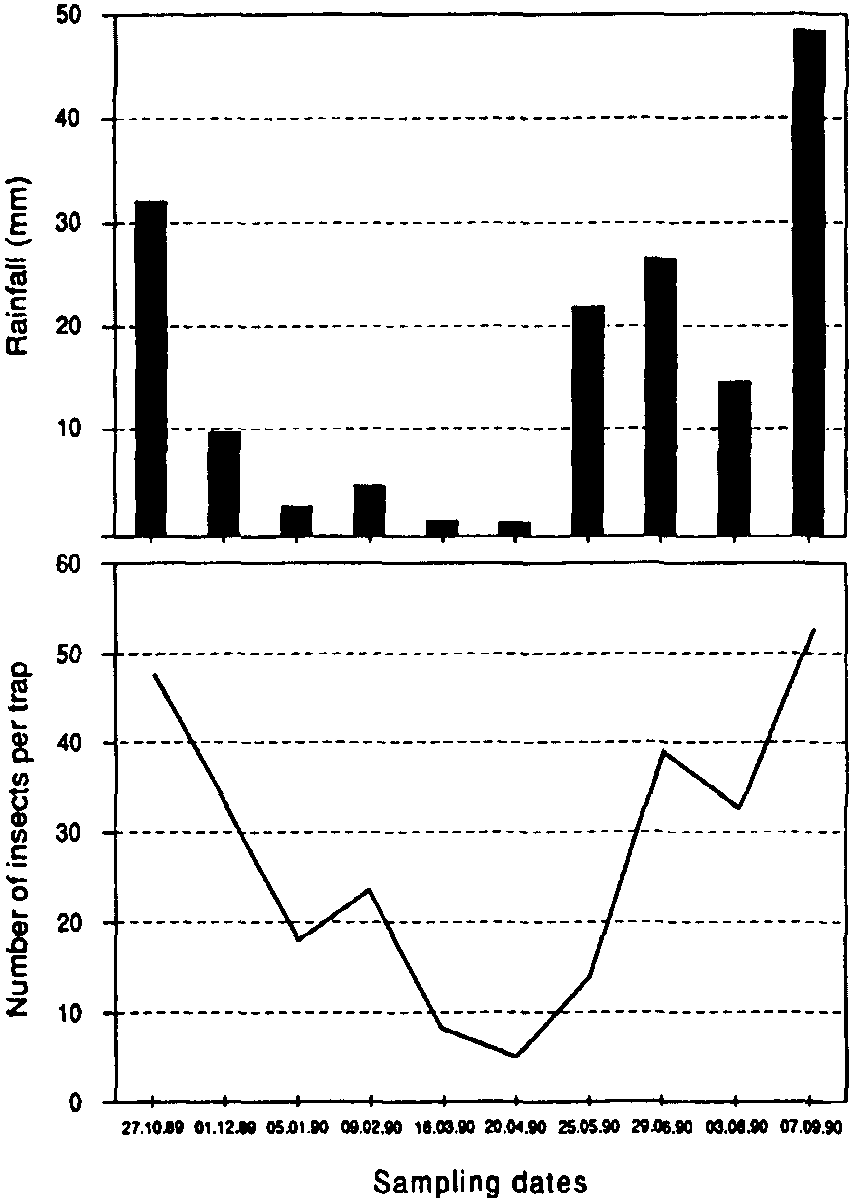
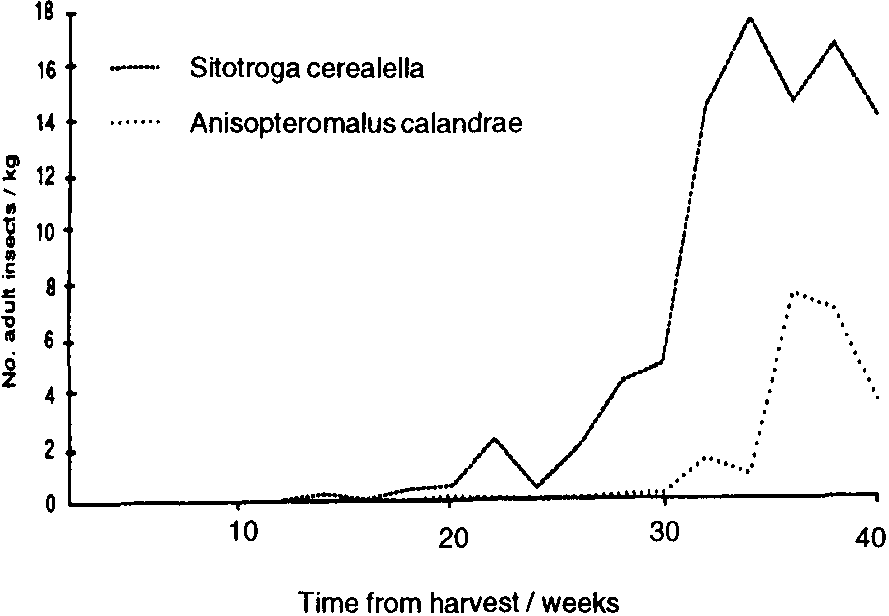
In Mexico, populations of all three primary pests remained low during the first three months of storage. Probably winter temperatures were low enough, on average, to delay population increase. Thereafter, populations of P. truncatus increased steadily; 40 weeks after harvest, populations of P. truncatus averaged 150-350 adults /kg in the three stores. with no indication that they were reaching their limit (Figure 1). Pheromone trap catches (Figure 2a) were high initially, low throughout the winter months and then increased sharply in May; unbaited sticky traps in the stores showed a similar pattern (Figure 2b). Data on emergences were too erratic (due to the clumped nature of P. truncatus oviposition) to allow conclusive evaluation of the relative importance of migration and reproduction to population build-up in store. However, a sharp increase in adult populations in one of the three experimental cribs after 36 weeks in store appeared to fallow a major increase in flight activity and so may have been due to immigration. Populations of the other two primary pests, Sitophilus ?zeamais and S. cerealella reached a «plateau» after 5-6 months, at a level of approximately 20 insects/kg (Figure 1). Damage to the maize was severe, weight losses reaching 16% after 42 weeks in store, and increased closely in line with the P. truncatus population (Figure 3).
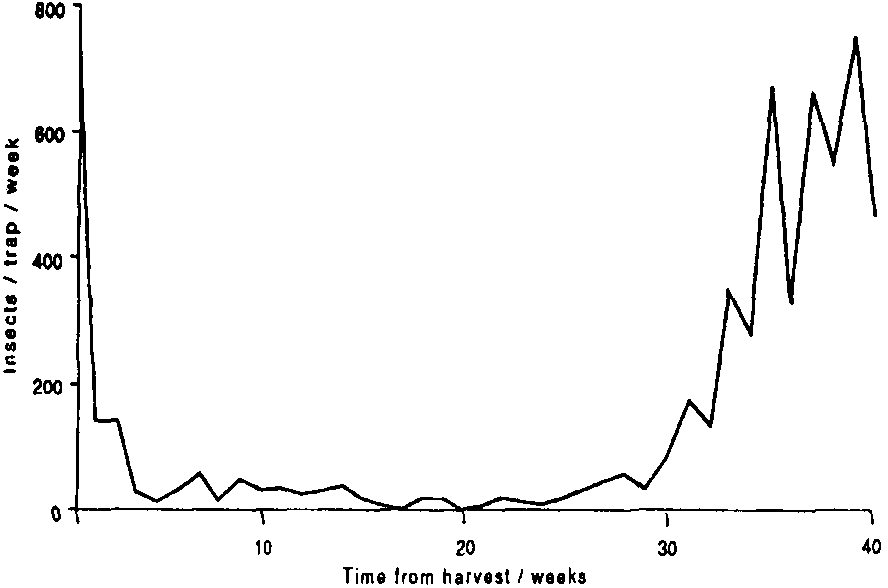
In Honduras, the pattern of infestation was quite different, with S. zeamais dominating the storage fauna and increasing, apparently exponentially, from the fourth month in store. P. truncatus was not recorded until the twelfth week in store, and then only a single specimen. Numbers began to increase and reproduction was detected in the stores after 21 weeks of storage. Significant infestation of the stores apparently followed a sharp increase in P. truncatus flight activity which accompanied the onset of the rainy season in May ( Figure 4). it remains to be seen whether P. truncatus populations will eventually reach damaging levels, despite the prior establishment of S. zeamais.
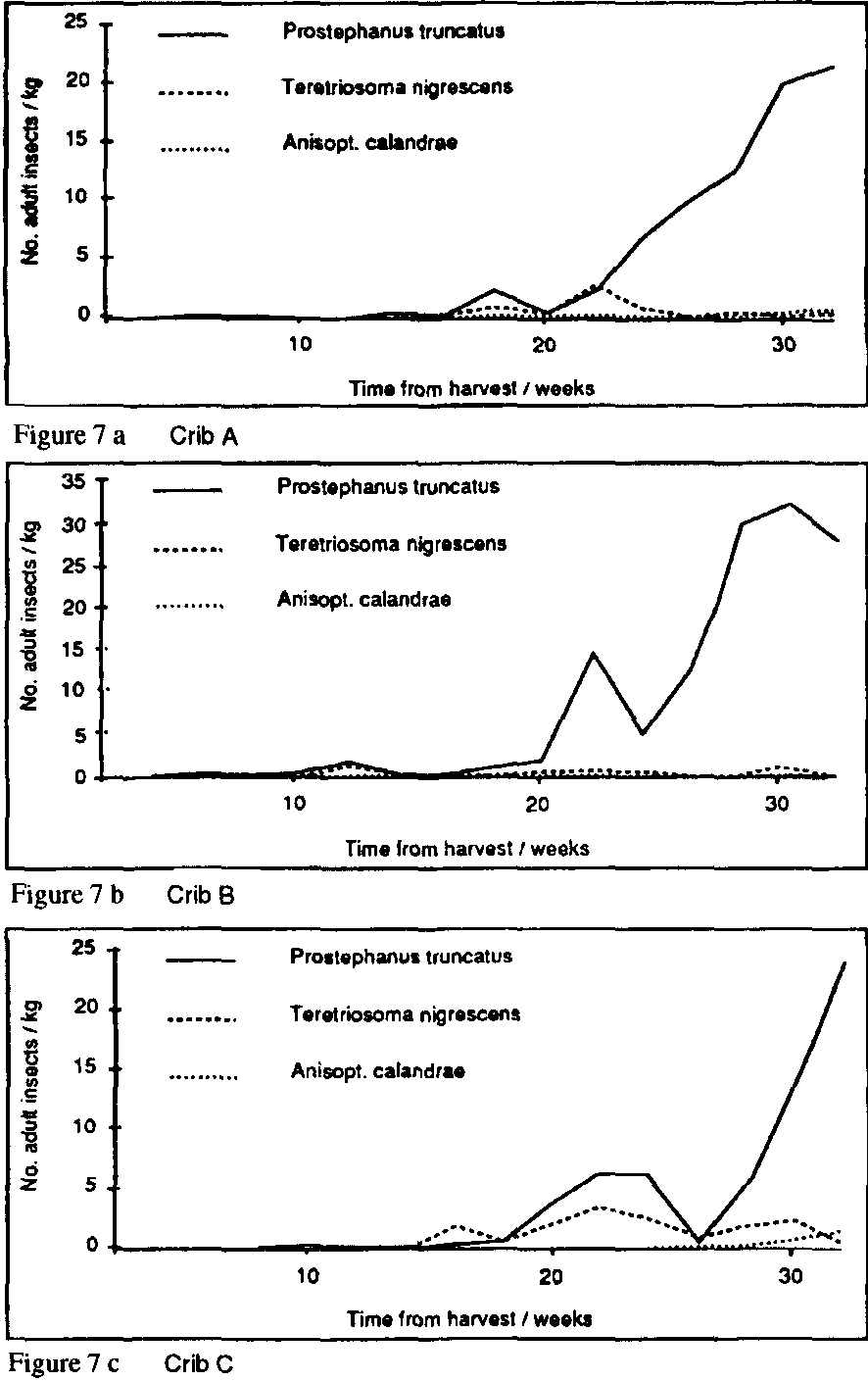
The predator T. nigrescens was found in flight traps at Chilcuautla throughout the period of observation (Figure 5) and was recorded in stores from the second sampling onwards (Figure 6). Adult populations in store fluctuated periodically, apparently reflecting immigration/emigration rather than reproduction/ mortality, though there was not a close correlation with «peaks,» in flight activity. There was no evidence at all that T. nigrescens could contain P. truncatus populations or that pest and predator reach an equilibrium, as reported earlier in Costa Rica (Boeye, 1988). indeed, T. nigrescens showed no numerical response whatsoever to the main P. truncatus population increase (Figure 7 a-c). There was some evidence that populations of the parasitoid Anisopteromalus calandrae (Hym.: Pteromalidae) were correlated with those of S. cerealella (Figure 8); however, this possible association cannot be properly evaluated without better data on emergences and more information on specificity. Certainly this parasitoid, recorded as a parasite of P. truncatus in Costa Rica (Boeye, 1988), showed no response to this host here (Figure 7). T. nigrescens has not yet been recorded in the experimental stores in Honduras nor in nearby flight traps, though small numbers have been recorded in traps elsewhere.
Figure 1 Build-up of major primary pests in stores (mean of 3 stores, 60 cobs per store)
Figure 2a. Captures of Prostephanus truncatus in pheromone traps, Chilcuautla,
Figure 2b. Captures of P. truncatus and T. nigrescens in unbaited sticky traps, Chilcuautla.
DISCUSSIONS AND CONCLUSIONS
Conclusions drawn at this time must be regarded as provisional, pending completion of data collection and processing. However, the situation at Chilcuautla where extremly destructive populations of P. truncatus develop in rural maize stores, despite the presence of coadapted natural enemies, clearly demonstrates the need for a cautious approach to biological control of this pest.
There is strong evidence in the data collected so far, that flight activity and immigration of the pest from populations outside the maize production and storage system may be crucial factors affecting the level of infestation. However, no specific source of infestation, in the form of an alternative host plant, has yet been identified in this study. There is also strong evidence that the annual cycle of P. truncatus flight activity is heavily dependent on weather, from the Honduras data, specifically on rainfall. Maize standing in the field, though it may become somewhat infested, does not appear to be highly attractive to dispersing P. truncatus It may even be that maize, as such, is not particularly attractive to P. truncatus and that maize stores become infested, to some extent "by accident", or because of their wooden structures. Once infested however, maize is clearly an excellent substrate for development and the aggregation pheromone ensures that initial colonisation is reinforced by further immigration. The implication would be that the timing of harvest, in relation to periods of flight activity, might play a very important part in determining the level of damage suffered by a particular store of maize. Stores which are filled well after the onset of dry weather (as is the case in the Mexican tropics and much of Central America) would tend not to become significantly infested until the following rainy season and would thus escape serious damage.
Major unknowns in such a «farming system,» interpretation of the situation remain the source of populations arriving from outside the maize production/storage system and in the relative importance of P. truncatus populations surviving in the structure of previously-infested stores in initiating early infestation of the newly-stored crop. It has been reported that a change in humidity stimulates P. truncatus to leave resting sites in wood (Detmers, 1988). it is not clear whether the addition of maize to an empty store, even in the dry season, would have the same effect. If early storage of maize, during periods of high humidity and high flight activity of P. truncatus does indeed lead to severe pest infestation, this has serious implications for management of the pest in Africa. In many areas, the severity of attack by lepidopterous stem borers on late-planted maize confines the production of maize to the early part of the rainy season (or the first rainy season in areas with a bimodal rainfall pattern); such maize must inevitably be harvested under humid conditions and may thus be exposed to early attack by P. truncatus in store.
Variation in the biotic potential of different populations of the pest and its natural enemies could be crucial factors determining the rate of pest population increase in an infested store and thus in determining whether an acceptable level of biological control can be achieved by natural enemies in a particular situation.
Research elsewhere has provided evidence that such variation exists, at least in this pest (Nissen, 1989), but this topic has yet to be investigated systematically. This will be a major target for investigations by BCP and its collaborators in 1990-91. Finally, the possible effect of different substrates, and particularly different maize varieties, on the biotic potential of the pest has also not been investigated properly; this too could critically affect the relationship between the pest and its natural enemies.
The complexity of the P. truncatus situation, already demonstrated by these preliminary studies, underlines the need for a holistic approach to management of this pest. The observations reported here do not necessarily imply that classical biological control will not he effective against P. truncatus. However, they do suggest that the introduction of T. nigrescens alone may well be insufficient to bring the pest under adequate control. Proper, quantitative evaluation is needed of the influence of natural enemies in relation to other factors determining pest population levels. Such evaluation can probably only be achieved through simulation modelling. The collection of the necessary data and the development of an appropriate model for this system should therefore be given the highest priority.
REFERENCES
Boeye, J. (1988) Autökologische Untersuchungen zum Verhalten des Grossen Kornbohrers Prostephanus truncatus (Horn) (Col.: Bostrichidae) in Costa Rica. Dissertation, Institut für Phytopathologie der Christian-Albrechts-Universität zu Kiel, 195pp.
Detmers, H.-B. (1988) Untersuchungen zur biologischen Bedeutung des Holzes für den Grossen Kornbohrer Prostephanus truncatus (Bostrichidae). MSc thesis, Freie Universität, Berlin. 121 pp.
Leliveldt, B., & G.-A. Laborius (1990) Effectiveness and specificity of the antagonist Teretriosoma nigrescens Lewis (Col.: Histeridae) on the Larger Grain Borer (Horn) (Col.: Bostrichidae). pp.87- 102 in R.H. Markham & H.R. Herren (Eds.) Biological control of the Larger Grain Borer. Proceedings of an IITA/ FAO coordination meeting, Cotonou, Republic of Benin, 2-3 June 1989, IITA, Ibadan, Nigeria, 167pp.
Markham, R.H. (1990) The role of quantitative ecology and systems analysis in the biological control of larger grain borer. pp 3550 in R.H. Markham & H.R. Herren (Eds.) Biological control of the Larger Grain Borer. Proceedings of an IITA/FAO coordination meeting, Cotonou, Republic of Benin, 2-3 June 1989, IITA, Ibadan, Nigeria, 167pp.
Markham &H.R. Herren (Eds.) (1990) Biological control of the Larger Grain Borer. Proceedings of an IITA/FAO coordination meeting, Cotonou, Republic of Benin, 2-3 June 1989, IITA, Ibadan, Nigeria, viii+167pp.
Nissen, U. (1989) Populationsvergleich von Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) unterschiedlicher geographischer Herkunft. MSc Thesis, Christian-Albrechts-Universität zu Kiel. 93pp.
Rees, D.P. (1987) Laboratory studies on predation by Teretriosoma nigrescens Lewis (Coleoptera: Histeridae) on Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) infesting maize cobs in the presence of other maize pests. J. Stored Prod. Res. 23, 191-195.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
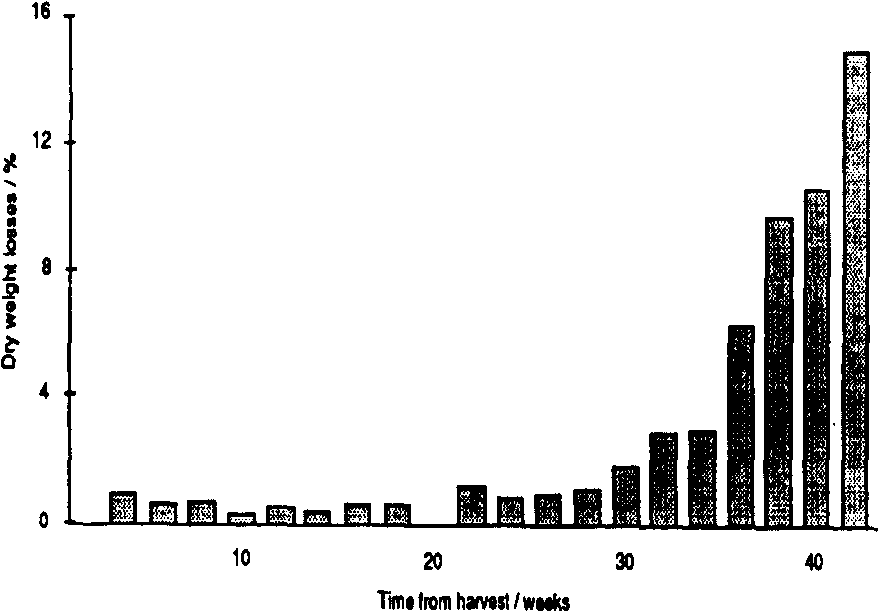{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}